

Keterangan
Kit Deteksi Mikoplasma GMyc-PCR terutama menggunakan metode PCR untuk mendeteksi infeksi Mikoplasma pada berbagai bahan biologis (seperti kultur sel, sekresi hewan percobaan, serum hewan, dll.). Kit ini menggabungkan beberapa keunggulan: sensitif, spesifik, cepat, dan dapat dideteksi secara langsung dengan supernatan kultur sel. Produk ini mendeteksi mikoplasma dalam bahan biologis seperti sel yang dikultur dengan metode PCR. Primer yang digunakan dirancang sesuai dengan wilayah yang dilestarikan dari urutan rRNA 16S-23S mikoplasma, dan hanya secara khusus mengamplifikasi DNA mikoplasma, dengan sensitivitas dan spesifisitas deteksi yang tinggi. Amplifikasi PCR dan analisis elektroforesis hanya membutuhkan waktu beberapa jam, dan operasinya mudah dan sederhana.
Kultur sel merupakan eksperimen umum dalam penelitian ilmu hayati. Tidak seperti metode eksperimen lain yang umum digunakan, kultur sel merupakan proses berkelanjutan yang dinamis, dan sel sering kali merespons kesalahan manipulasi atau kontaminan yang sering kali menunjukkan keadaan sel atau tampilan medium yang tidak normal. Jika terkontaminasi oleh mikoplasma, morfologi sel tidak mengalami perubahan yang jelas, dan mudah diabaikan. Sering kali tidak ditemukan hingga pencemarannya sangat serius. Mungkin terdapat ratusan mikoplasma pada membran sel yang terkontaminasi, mikoplasma ini bersaing untuk mendapatkan nutrisi dan melepaskan metabolit toksik, yang secara serius memengaruhi hasil eksperimen.
Penelitian telah menunjukkan bahwa setidaknya ada 20 jenis mikoplasma yang dapat mencemari sel, di antaranya yang paling umum adalah: Mikoplasma oral (M. orale), Mikoplasma arginin (M. arginin), Mikoplasma hyorhinis (M. hyorhini), Mikoplasma fermentasi (M. fermentans), Mikoplasma hominis (M. hominis), Mikoplasma salivarius (M.salivarium), Mikoplasma paru-paru (M. pulmonalis) dan Mycoplasma pir (M. pirum). Tingkat kontaminasi mikoplasma pada sel yang dikultur berkisar antara 4% hingga 92%. Sumber kontaminasi meliputi lingkungan kerja, operator itu sendiri (beberapa mikoplasma merupakan flora normal tubuh manusia), media kultur, serum, kontaminasi silang sel, peralatan eksperimen, dan kontaminasi yang digunakan pada jaringan atau organ asli tempat sel disiapkan.
Mengidentifikasi penyebab utama masalah selama kultur sel merupakan tugas yang sulit dan memakan waktu, di mana perubahan mendadak apa pun harus dicurigai, dan praktik pengujian yang baik serta pengujian rutin untuk kontaminasi mikoplasma diperlukan. Ada banyak metode untuk mendeteksi mikoplasma, seperti kultur langsung, pewarnaan fluoresensi DNA, ELISA, dan metode PCR.
Fitur
- Primer yang digunakan dirancang berdasarkan daerah konservasi sekuens rRNA Mycoplasma 16S-23S.
- Hanya DNA mikoplasma yang diperkuat secara khusus
- Sensitivitas dan spesifisitas yang sangat baik
- Lebih dari 20 spesies mikoplasma dapat dideteksi
Aplikasi
- Deteksi Mikoplasma
Komponen
| Komponen No. | Nama | 40601ES10 (10 pengujian) | 40601ES20 (20 pengujian) |
| 40601-A | Campuran PCR GMyc-1 | 250 µL | 2×250 µL |
| 40601-B | Campuran PCR GMyc-2 | 250 µL | 2×250 µL |
| 40601-C | Template Kontrol Positif N | 20 µL | 20 µL |
[Catatan] 1. Bila tidak digunakan dalam waktu lama, dapat disimpan dalam keadaan beku pada suhu -85~-65℃.
2. Reaksi PCR sangat sensitif.Untuk mencegah positif palsu, kontrol positif ditambahkan di akhir saat menambahkan sampel.
Penyimpanan
Produk ini dapat disimpan pada suhu -25~-15℃ selama 18 bulan. Jika tidak digunakan dalam waktu lama, harap jauhkan dari cahaya.
Angka
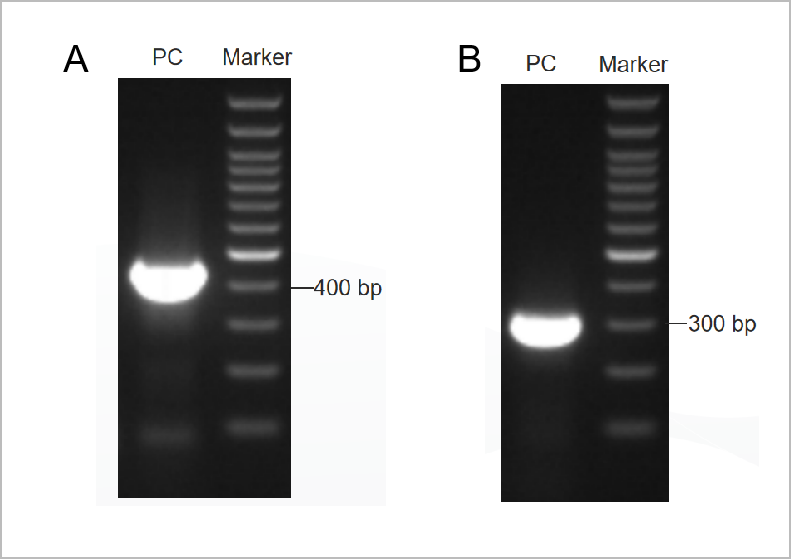
Gambar 1. Hasil elektroforegram PCR Mycoplama Test Kit.
Elektroforetogram putaran pertama ditunjukkan pada Gambar 1A dan elektroforetogram putaran kedua ditunjukkan pada Gambar 1B. (M: penanda 1kb, 1-2: kontrol positif, 3: kontrol negatif)
[1] Rao XS, Cong XX, Gao XK, dkk. Fosforilasi yang dimediasi AMPK meningkatkan penghambatan otomatis TBC1D17 untuk meningkatkan penyerapan glukosa yang bergantung pada Rab5. Cell Death Differ. 2021;28(12):3214-3234. doi:10.1038/s41418-021-00809-9(IF:15.828)
[2] Guo F, Li L, Li J, dkk. Pengurutan multiomik sel tunggal embrio awal tikus dan sel induk embrionik. Cell Res. 2017;27(8):967-988. doi:10.1038/cr.2017.82(IF:15.606)
[3] Hao Y, He B, Wu L, dkk. Translokasi nuklir p85β meningkatkan tumorigenesis kanker mutan domain heliks PIK3CA. Nat Commun. 2022;13(1):1974. Diterbitkan 13 April 2022. doi:10.1038/s41467-022-29585-x(IF:14.919)
[4] Shu X, Liu M, Lu Z, dkk. Pemetaan genom secara luas menunjukkan bahwa deoksiuridin diperkaya dalam DNA sentromerik manusia. Nat Chem Biol. 2018;14(7):680-687. doi:10.1038/s41589-018-0065-9(IF:13.843)
[5] Li X, Xiong X, Wang K, dkk. Pemetaan di seluruh transkriptom mengungkap metilom N(1)-metiladenosin yang reversibel dan dinamis. Nat Chem Biol. 2016;12(5):311-316. doi:10.1038/nchembio.2040(IF:12.709)
[6] Sun L, Yang X, Huang X, dkk. 2-Hidroksilasi Asam Lemak Menekan Tumorigenesis dan Metastasis Kolorektal melalui Sumbu Transkripsi YAP. Cancer Res. 2021;81(2):289-302. doi:10.1158/0008-5472.CAN-20-1517(IF:12.701)
[7] Sun Z, Zhang Z, Wang QQ, Liu JL. Inaktivasi Gabungan CTPS1 dan ATR Secara Sintetis Mematikan Sel Kanker yang Mengekspresikan MYC Secara Berlebihan. Cancer Res. 2022;82(6):1013-1024. doi:10.1158/0008-5472.CAN-21-1707(IF:12.701)
[8] Song J, Zhuang Y, Zhu C, dkk. Peran diferensial PUS10 manusia dalam pemrosesan miRNA dan pseudouridilasi tRNA. Nat Chem Biol. 2020;16(2):160-169. doi:10.1038/s41589-019-0420-5(IF:12.154)
[9] He B, Pan H, Zheng F, dkk. RNA noncoding panjang LINC00930 meningkatkan glikolisis tumor dan proliferasi sel yang dimediasi PFKFB3 pada karsinoma nasofaring. J Exp Clin Cancer Res. 2022;41(1):77. Diterbitkan 24 Februari 2022. doi:10.1186/s13046-022-02282-9(IF:11.161)
[10] Tang B, Liu BH, Liu ZY, Luo MY, Shi XH, Pang DW. Titik Kuantum dengan Lapisan Zwitterionik Amfifilik Kompak. Antarmuka Aplikasi ACS. 2022;14(24):28097-28104. doi:10.1021/acsami.2c04438(IF:9.229)
[11] Huang C, Zhang Z, Chen L, dkk.Asetilasi dalam Domain N- dan C-Terminal Src Mengatur Peran Berbeda dari Tumorigenesis yang Dimediasi STAT3. Cancer Res. 2018;78(11):2825-2838. doi:10.1158/0008-5472.CAN-17-2314(IF:9.130)
[12] Wu X, Yu M, Zhang Z, dkk. DDB2 mengatur replikasi DNA melalui degradasi CDT2 yang independen dari PCNA. Cell Biosci. 2021;11(1):34. Diterbitkan 8 Februari 2021. doi:10.1186/s13578-021-00540-5(IF:7.133)
[13] Wang J, Zhang Y, Liu X, Liu H. Mengoptimalkan Terapi Adaptif Berdasarkan Keterjangkauan Subpopulasi yang Tahan Tumor. Cancers (Basel). 2021;13(21):5262. Diterbitkan 20 Oktober 2021. doi:10.3390/cancers13215262(IF:6.639)
[14] Feng W, Liu R, Xie X, dkk. SUMOylation dari α-tubulin adalah modifikasi baru yang mengatur dinamika mikrotubulus. J Mol Cell Biol. 2021;13(2):91-103. doi:10.1093/jmcb/mjaa076(IF:6.216)
[15] Yu M, Hu X, Yan J, Wang Y, Lu F, Chang J. Inhibitor RIOK2 NSC139021 Memberikan Efek Anti-Tumor pada Glioblastoma melalui Induksi Penghentian Siklus Sel dan Apoptosis yang Dimediasi Skp2. Biomedicines. 2021;9(9):1244. Diterbitkan 17 September 2021. doi:10.3390/biomedicines9091244(IF:6.081)
[16] Ren S, Cai Y, Hu S, dkk. Berberin memberikan aktivitas antitumor pada limfoma sel B besar difus dengan memodulasi sumbu c-myc/CD47. Biochem Pharmacol. 2021;188:114576. doi:10.1016/j.bcp.2021.114576(IF:5.858)
[17] Wen F, Sun X, Sun C, dkk. TAGLN Diturunkan oleh Degradasi Proteasomal yang Dimediasi TRAF6 pada Sel Kanker Prostat. Mol Cancer Res. 2021;19(7):1113-1122. doi:10.1158/1541-7786.MCR-20-0513(IF:5.852)
[18] Tang B, Sun EZ, Zhang ZL, dkk. Domain Kolesterol yang Disekuestrian dengan Sphingomyelin Merekrut Protein Pengikat Formin 17 untuk Menyempitkan Lubang Berlapis Clathrin pada Masuknya Virus Influenza. J Virol. 2022;96(5):e0181321. doi:10.1128/JVI.01813-21(IF:5.103)
[19] Hu J, Ren W, Qiu W, dkk. Pembuatan garis sel punca pluripoten terinduksi (XDCMHi001-A) dari pasien Ankylosing spondylitis dengan mutasi JAK2. Stem Cell Res. 2020;45:101788. doi:10.1016/j.scr.2020.101788(IF:4.495)
[20] Xiao S, Yao X, Ye J, Tian X, Yin Z, Zhou L. Modifikasi epigenetik memfasilitasi ekspresi abnormal prolin sintase PYCR1 pada kanker lambung [diterbitkan online sebelum cetak, 30 Mei 2022]. Biochim Biophys Acta Gene Regul Mech. 2022;1865(6):194829. doi:10.1016/j.bbagrm.2022.194829(IF:4.490)
[21] Wang J, Zhang Y, Liu X, Liu H. Apakah Perawatan Periodik Tetap Efektif untuk Sistem Tumor Tanpa Informasi Lengkap?. Cancer Manag Res. 2021;13:8915-8928. Diterbitkan 30 November 2021. doi:10.2147/CMAR.S339787(IF:3.989)
[22] Yang X, Ren S, Rehman ZU, dkk. Karakterisasi molekuler, ekspresi, dan identifikasi fungsional kinase pengikat TANK 1 (TBK1) pada sapi (Bos taurus) dan kambing (Capra hircus). Dev Comp Immunol. 2022;133:104444. doi:10.1016/j.dci.2022.104444(IF:3.636)
[23] Zheng D, Chang X, Liu Y, dkk. 2-Methoxy-5((3,4,5-trimethosyphenyl)seleninyl) fenol membalikkan migrasi dan invasi sel yang diinduksi EGF melalui penurunan regulasi MDM2 pada lini sel kanker payudara. Cancer Biol Ther. 2019;20(4):513-523. doi:10.1080/15384047.2018.1537578(IF:3.373)
[24] Xu F, Zhang S, Liu Z, dkk. TEX9 dan eIF3b bersinergi secara fungsional untuk meningkatkan perkembangan karsinoma sel skuamosa esofagus. BMC Cancer. 2019;19(1):875. Diterbitkan 3 September 2019. doi:10.1186/s12885-019-6071-9(IF:2.933)
[25] Pan H, Sun L, Wang W, dkk. Serum long non-coding RNA LOC553103 sebagai biomarker diagnostik dan prognostik non-spesifik untuk jenis kanker manusia yang umum. Clin Chim Acta. 2020;508:69-76. doi:10.1016/j.cca.2020.05.017(IF:2.615)
[26] Li N, Lin SM, Li Y, Sun J, Zhang L, Chen M.Garis sel punca pluripoten terinduksi (GZHMCi004-A) yang berasal dari janin dengan mutasi heterozigot G380R pada gen FGFR3 yang menyebabkan akondroplasia. Stem Cell Res. 2021;53:102322. doi:10.1016/j.scr.2021.102322(IF:2.020)
[27] Luo Q, Wei C, Long Y, dkk. Pembuatan garis sel punca embrionik manusia knockout ELTD1 melalui sistem iCRISPR/Cas9. Stem Cell Res. 2021;53:102350. doi:10.1016/j.scr.2021.102350(IF:2.020)
[28] Liu YQ, Ling TW, Wang HY, Yang YH, Song WJ, Wang TC. Pembuatan garis sel punca pluripoten terinduksi bebas integrasi (LZUSHI001-A) dari pasien epilepsi dengan mutasi DGKG. Stem Cell Res. 2022;61:102768. doi:10.1016/j.scr.2022.102768(IF:2.020)
[29] Chen M, Lin SM, Li N, Li Y, Li Y, Zhang L. Garis sel punca pluripoten terinduksi (GZHMCi003-A) yang berasal dari janin dengan delesi heterozigot ekson 3 pada gen RUNX2 yang menyebabkan displasia kleidokranial. Stem Cell Res. 2021;51:102166. doi:10.1016/j.scr.2021.102166(IF:2.020)
[30] Xu Y, Wang X, Qiu T, et al. Pembuatan garis sel induk pluripoten yang diinduksi (FDCHI007-A) yang berasal dari pasien dengan ensefalopati perkembangan dan epilepsi Tipe 31 yang membawa c.545C heterozigot > Mutasi pada gen DNM1. Stem Cell Res. 2022;60:102709. doi:10.1016/j.scr.2022.102709(IF:2.020)
[31] Fan T, He J, Wang Y, Yu J, Sun W. Pembuatan garis sel punca pluripoten terinduksi (FDCHi006-A) dari seorang gadis berusia 7 tahun dengan pubertas prekoks sentral. Stem Cell Res. 2021;56:102542. doi:10.1016/j.scr.2021.102542(IF:2.020)
[32] Gong X, Zheng Z, Yang T, Zheng H, Xiao X, Jia N. Pembuatan garis keturunan iPSC yang dikoreksi gen secara isogenik (OGHFUi001-A-1) dari pasien ensefalopati epilepsi infantil dini tipe 1 (EIEE1) dengan mutasi hemizigot R330L pada gen ARX. Stem Cell Res. 2022;60:102693. doi:10.1016/j.scr.2022.102693(IF:2.020)
[33] Jia N, Gong X, Chen J, dkk. Pembuatan garis sel punca pluripoten terinduksi (OGHFUi001-A) dari ensefalopati epilepsi infantil dini tipe 1 dengan mutasi ARX. Stem Cell Res. 2021;53:102367. doi:10.1016/j.scr.2021.102367(IF:2.020)
[34] Zhu W, Zhou Y, Wang Q, dkk. Pembuatan garis sel punca pluripoten terinduksi (iPSC) manusia dari fibroblas kulit pasien yang membawa mutasi E363Q pada gen PSEN1. Stem Cell Res. 2022;61:102769. doi:10.1016/j.scr.2022.102769(IF:2.020)
[35] Luo F, Long K, Li X, dkk. Kekurangan LRRC8A melemahkan nekrosis yang disebabkan oleh hipoksia pada sel 3T3-L1. Biosci Biotechnol Biochem. 2020;84(6):1139-1145. doi:10.1080/09168451.2020.1730689(IF:1.516)
Pembayaran & Keamanan
Informasi pembayaran Anda diproses dengan aman. Kami tidak menyimpan detail kartu kredit atau memiliki akses ke informasi kartu kredit Anda.
Pertanyaan
Anda mungkin juga menyukai
FAQ
Produk ini hanya untuk keperluan penelitian dan tidak ditujukan untuk penggunaan terapeutik atau diagnostik pada manusia atau hewan. Produk dan konten dilindungi oleh paten, merek dagang, dan hak cipta milik
Aplikasi tertentu mungkin memerlukan hak kekayaan intelektual pihak ketiga tambahan.