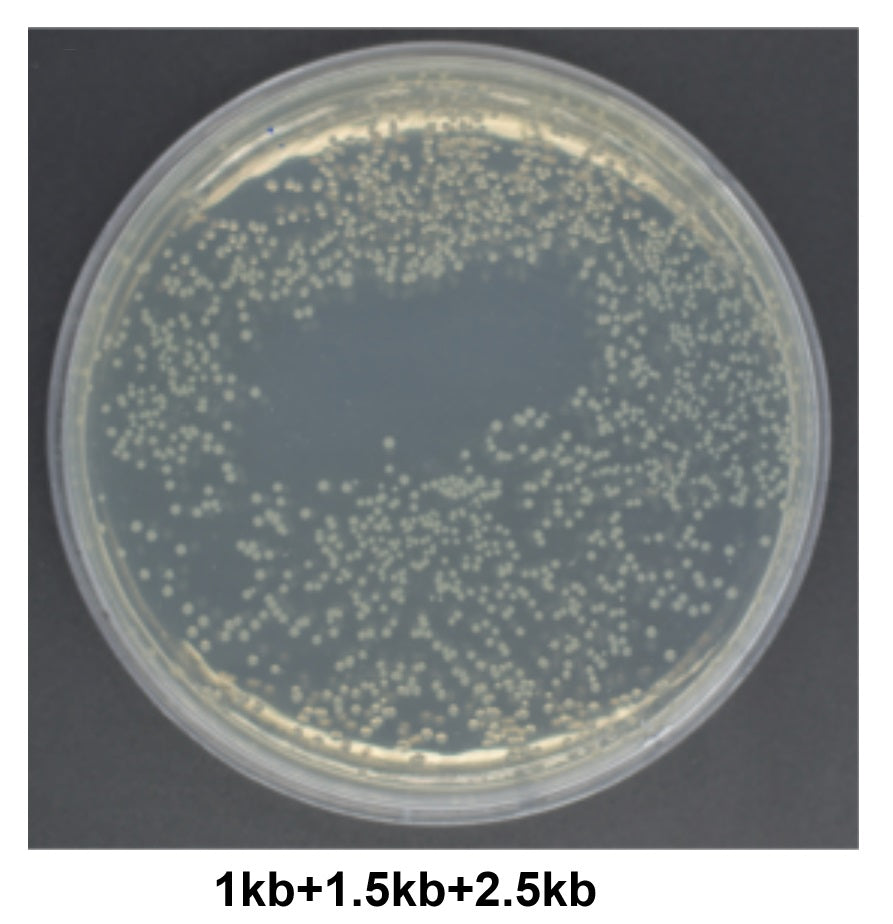
図 3. 超長断片のライゲーション。結果は、10923ES のコロニー数が多く、陽性率が 100% であることを示しています。A: 組換え形質転換プレート。ベクター (21 kb) と挿入断片 (4.5 kb) のモル比は 1:2 です。B: 挿入断片の PCR 同定電気泳動、M: Yeasen 10505ES。ベクター量: 20 μL 反応システムで 200 ng、反応条件: 50°C、40 分。


説明
Hieff Clone™ Universal II One Step Cloning Kit は、新しくアップグレードされたクローニング キットです。
慎重に最適化された第2世代2×Hieff Clone™ Universal II Enzyme Premix は、組換え酵素と組換え反応に必要なバッファーを組み合わせ、独自の組換えエンハンサーを追加することで、組換えクローニングの効率を大幅に向上させます。
このキットは、PCR産物を任意のベクターの任意のサイトにクローニングするように指示することができ、未精製PCR産物、直接回収されたPCR産物、低濃度のゴム回収産物と互換性があり、この製品は、結合断片の30%〜70%の相同アームGC含有量を再構成することができます。ベクターは完全に線状化され、線状化ベクターの末端の15〜25 bpの相同配列が、挿入された断片の陽性および逆PCRプライマーの5 '末端に導入されたため、挿入された断片のPCR産物の5 '末端と3 '末端は、それぞれ線状化ベクターの両端に対応するまったく同じ配列を持ちました。組み換え酵素の作用により、PCR産物と線状化ベクターの組み換え反応は、50℃でわずか5分で完了できます。クローニングの陽性率は95%以上に達することができます。
特徴
シンプル: 1 回の反応で最大 6 個の DNA 断片をクローン化します。
柔軟性: 設計ガイドラインにより、任意のベクトルに組み立てることができます。
効率的: 1 ~ 7 個の断片の連結に効率的です。
アプリケーション
急速な クローニング: 複数の重複する DNA 断片を 1 ステップの 5 ~ 50 分の等温反応でシームレスにリンクできます。
指向性クローニング; 部位特異的突然変異誘発。
仕様
|
結紮 |
最大7個のフラグメント |
|
製品タイプ |
DNAアセンブリ |
コンポーネント
|
コンポーネント番号 |
名前 |
10923ES05 ( 5T ) |
10923ES20 ( 20T ) |
10923ES50 ( 50T ) |
|
10923 -A |
2 ×ハイフクローンTM ユニバーサル II 酵素プレミックス |
50μL |
200μL |
500μL |
|
10923 -B |
500 bp コントロールインサート ( 2 5 ng/ μL ) |
5μL |
5μL |
5μL |
|
10923 -C |
pUC 19コントロールベクター、線形化(50 ng/ μL ) |
5μL |
5μL |
5μL |
ストレージ
この製品は-25〜-15℃で1年間保管してください。 年。
数字

形 1. 10 ngの低入力単一フラグメントを、6μLの小型システムの非蘇生受容状態で接続しました。 結果は、10923ES が競合製品よりも優れた性能を示し、接続効率が高く、コロニーが多く、陽性率が 100% であることを示しました。AB: クローンプレート。キャリア (10 kb) と挿入断片 (1 kb) のモル比は 1:2 でした。C: 挿入断片 PCR によって識別された電気泳動マップ、M: YEASEN 10505ES。

図2. 6つの断片(全長7.6 kb)を11.6 kbプラスミドにクローニングしました。結果は、10923ESが競合製品よりも優れた性能を示し、コロニーが多く、陽性率が100%であることを示しました。AB:コロニープレート。キャリア(11.6 kb)と挿入断片のモル比は1:2でした。C:挿入断片の電気泳動分析、M:YEASEN 10505ES。プラスミドインプット: 200 ng/20 μL、50°C、50分。

関連する引用と参考文献:
[1] Zhang H, Shao S, Zeng Y, et al. 熱ショック時の急性転写応答にはHSF1の可逆的な相分離が必要である。Nat Cell Biol . 2022;24(3):340-352. (IF:28.824)
[2] Xu Y、Yu Q、Wang P、et al。選択的小分子c-Myc分解剤は致死的なc-Myc過剰発現腫瘍を強力に退縮させる。Adv Sci (Weinh)。2022;9(8):e2104344。 (IF:16.806)
[3] Guan B、Jiang YT、Lin DL、Lin WH、Xue HW。ホスファチジン酸はGAPC(グリセルアルデヒド-3-リン酸デヒドロゲナーゼ)およびPGK(ホスホグリセリン酸キナーゼ)タンパク質に結合することで競合阻害を介してオートファジーを抑制する[印刷に先駆けてオンライン出版、2022年3月15日]。オートファジー。2022;1-15。 (IF:16.016)
[4] He X、Li Y、Chen Q、et al。SIRT7のO-GlcNAcylationと安定化はSIRT7-REGγ相互作用を阻害することで膵臓癌の進行を促進する[印刷に先駆けてオンライン出版、2022年4月14日]。Cell Death Differ。2022 ;10.1038/s41418-022-00984-3。 (IF:15.828)
[5] Zhou T、Zhu X、Ye Z、et al。ループスエンハンサーリスクバリアントは、lncRNAとDNAメチル化機構の協調によりIRF8の調節異常を引き起こす。Nat Commun。2022 ;13(1):1855。2022年4月6日発行。 (IF:14.919)
[6] Li T、Chen X、Qian Y、et al。マウスのグルコース恒常性を可能にする脈動遺伝子発現のための合成BRETベースの光遺伝学デバイス。Nat Commun。2021 ;12(1):615。2021年1月27日発行。 (IF:14.919)
[7] Wu P、Zhang T、Liu B、et al. ペプチド-MHCクラスIコンフォメーションの機械的制御がTCR抗原認識を決定する。Mol Cell. 2019;73(5):1015-1027.e7. (IF:14.548)
[8] Xiong L、Liu S、Chen S、et al。古細菌における新しいタイプのDNAホスホロチオ化ベースの抗ウイルスシステム。Nat Commun。2019 ;10(1):1688。2019年4月11日発行。 (IF:11.878)
[ 9 ] Jiang L、Wang Y、Xia A、et al.亜硫酸還元酵素の天然一塩基多型変異はトウモロコシの硫黄同化に影響を与える。New Phytol. 2021;232(2):692-704. (IF:10.152)
支払いとセキュリティ
お支払い情報は安全に処理されます。 クレジットカードの詳細を保存したり、クレジットカード情報にアクセスすることはありません
問い合わせ
あなたも好きかもしれません
よくある質問
この製品は研究目的のみに使用され、人間や動物の治療や診断に使用することを意図したものではありません。製品とコンテンツは、
特定のアプリケーションでは、追加のサードパーティの知的財産権が必要になる場合があります。