





説明
Hieff Unicon™ユニバーサルブルーqPCRマスターミックス(染料ベース)は、高感度と特異性を特徴とする2×リアルタイム定量PCR増幅用のプレソリューションで、青色で、サンプル添加トレース効果があります。コアコンポーネントのHieff Unicon™Taq DNAポリメラーゼは、抗体ホットスタートを使用して、サンプル調製中のプライマーアニーリングによる非特異的増幅を効果的に抑制します。同時に、フォーミュラは促進因子を追加してPCR反応の増幅効率を向上させ、異なるGC含有量(30〜70%)の遺伝子の増幅を均等化し、定量PCRが広い定量領域で良好な線形関係を得ることができるようにします。この製品には、ほとんどのqPCR機器に適用できる特別なROXパッシブリファレンスダイが含まれています。異なる機器でROXの濃度を調整する必要はありません。プライマーとテンプレートを追加して、増幅用の反応システムを準備するだけです。
特徴
- 内蔵の視覚インジケーター: リアルタイム PCR を妨げず、チューブやウェル内での視認性を高める不活性な青色染料が含まれています。
- 高い特異性: Taq DNA ポリメラーゼが含まれており、抗体を介した熱プライミング機構によって厳密に制御されるため、非特異的な増幅を回避できます。
- 幅広い機器互換性:ほとんどのqPCR機器と互換性があり、異なる機器でROXの濃度を調整する必要がありません。
アプリケーション
- 遺伝子発現
仕様
| 集中 | 2× |
| 検出方法 | SYBRグリーン染料 |
| PCR法 | 定量PCR |
| ポリメラーゼ | Taq DNAポリメラーゼ |
| サンプルの種類 | DNA |
| アプリケーション機器 | ほとんどのqPCR機器 |
| 製品タイプ | リアルタイム蛍光定量PCR用プレミックス |
| 申請する(アプリケーション) | 遺伝子発現 |
コンポーネント
| コンポーネント番号 | 名前 | 11184ES03 | 11184ES08 | 11184ES50 | 11184ES60 |
| 11184 |
Hieff Unicon™ ユニバーサル ブルーqPCRマスター ミックス (染料ベース) |
1mL | 5×1mL | 50×1mL | 100×1mL |
配送と保管
製品は保冷剤とともに出荷され、-15℃~-25℃で18ヶ月間保存できます。製品には蛍光色素が含まれているため、保管時や反応系を調製する際には強い光の照射を避ける必要があります。
数字
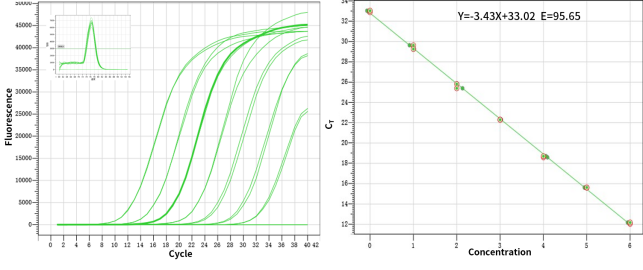
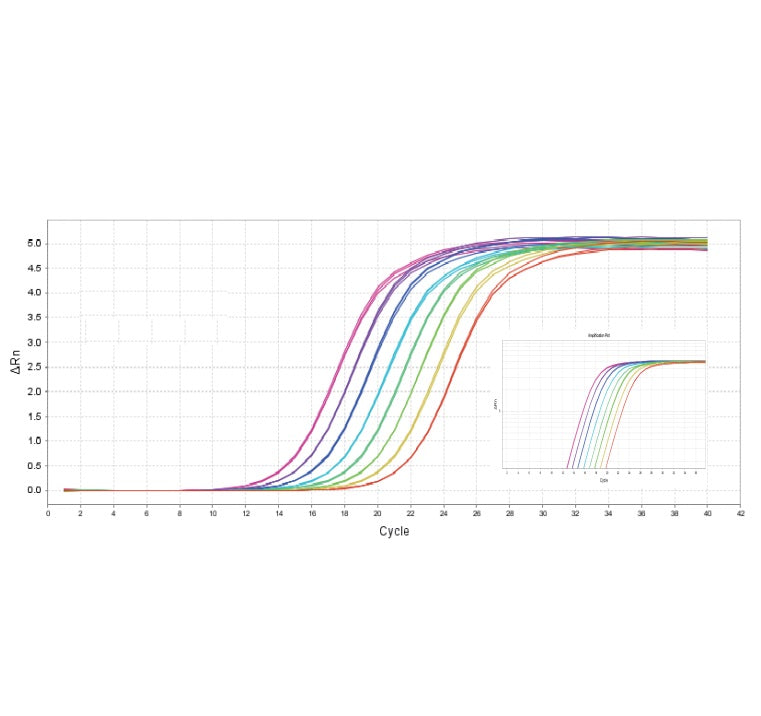
図 1. 高感度: 単一のコピーを検出する能力。
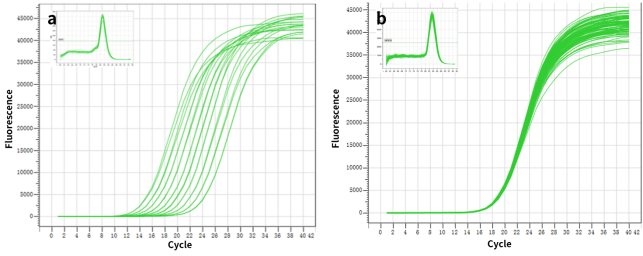
図2. 高解像度と重複ウェルの優れた再現性。
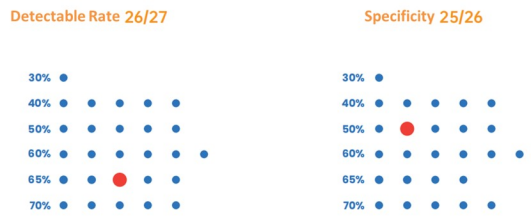
図3. 優れた検出率と特異性:30~70% GCの増幅に広く適しています。

「VCPはUSP2をリクルートしてFASNのユビキチン化と分解を阻害することでオートファジー関連の骨肉腫の進行を促進する」 、 Cell Death & Diseaseより引用。 第15巻、論文番号:788(2024)"

「SARS-CoV-2エンベロープタンパク質は急性呼吸窮迫症候群(ARDS)のような病理学的損傷を引き起こし、抗ウイルスの標的となる。Cell Res. 2021;31(8):847-860. doi:10.1038/s41422-021-00519-4」より引用

「形質膜と葉緑体を結び、病原体によって利用される防御経路。Cell . 2020年9月3日;182(5):1109-1124.e25. doi: 10.1016/j.cell.2020.07.020.」より引用
[1] Xia B、Shen X、He Y、et al。SARS-CoV-2エンベロープタンパク質は急性呼吸窮迫症候群(ARDS)のような病理学的損傷を引き起こし、抗ウイルスの標的となる。Cell Res. 2021;31(8):847-860. doi:10.1038/s41422-021-00519-4(IF:25.617)
[2] Wang S、Li Y、Zhong L、et al。細胞内のイントロン選択マーカーを介した効率的な遺伝子編集。Cell Mol Life Sci。2022;79(2):111。2022年1月31日発行。doi:10.1007/s00018-022-04152-1(IF:9.261)
[3] An LL、Zhao X、Gong XY、et al。IFN応答におけるゼブラフィッシュIRFファミリーメンバーのプロモーター結合および核保持特性。Front Immunol。2022;13:861262。2022年4月6日発行。doi:10.3389/fimmu.2022.861262(IF:7.561)
[4] Zhao Y、Wang HP、Yu C、et al。誘導ストレス条件下でのHaematococcus pluvialisにおけるメラトニンのアスタキサンチンと脂質の共生産に対する刺激効果の背後にある制御メカニズムを解明するための生理学的および代謝学的プロファイルの統合。Bioresour Technol。2021;319:124150。doi:10.1016 / j.biortech.2020.124150(IF:7.539)
[5] Shu C、Wang L、Zou C、et al. ヒラメParalichthys olivaceusの生殖腺分化におけるFoxl2およびDmrt1タンパク質の機能[印刷に先駆けてオンライン出版、2022年6月16日]。Int J Biol Macromol. 2022;215:141-154. doi:10.1016/j.ijbiomac.2022.06.098(IF:6.953)
[6] Wang S、Huang J、Liu F、et al。イソステビオールナトリウムは、代謝再プログラミングと免疫応答調節を介して、デキストラン硫酸ナトリウム誘発性大腸炎を伴うBALB/cマウスに抗大腸炎効果を発揮します。J Inflamm Res. 2021;14:7107-7130。2021年12月20日発行。doi:10.2147/JIR.S344990(IF:6.922)
[7] Wang J、Hu R、Wang Z、et al。レンチウイルス媒介SV40TおよびhTERT遺伝子導入による不死化ヤクルーメン上皮細胞株の確立。Oxid Med Cell Longev。2022;2022:8128028。2022年3月25日発行。doi:10.1155/2022/8128028(IF:6.543)
[8] Liu W、Guan Y、Qiao S、et al。Vicatia thibetica de Boiss根エキスのCaenorhabditis elegansおよび成体マウスのドキソルビシン誘発性早期老化に対する抗老化効果。Oxid Med Cell Longev。2021;2021:9942090。2021年8月6日発行。doi:10.1155/2021/9942090(IF:6.543)
[9] Qian Z、Liu C、Li H、et al。オステオカルシンはマクロファージにおけるGPR37の活性化を介してリポ多糖誘発性急性炎症を軽減する。バイオメディシンズ。2022;10(5):1006。2022年4月27日発行。doi:10.3390/biomedicines10051006(IF:6.081)
[10] Zhao X、Huang Y、Li X、et al. キトサン修飾マイクロスフェアを用いた遠心マイクロ流体チップへの核酸抽出および検出の完全統合 [印刷前にオンラインで公開、2022年6月27日]。Talanta。2022;250:123711。doi:10.1016/j.talanta.2022.123711(IF:6.057)
[11] Xu X、Wang H、Li X、Duan X、Wang Y。新しいALG10/TGF-β正の調節ループが大腸癌の幹細胞性に寄与する。Aging(Albany NY)。2022;14(11):4858-4873。doi:10.18632/aging.204116(IF:5.955)
[12] Li F、Hu X、Wu J. Daidzein は Clec11a のアップレギュレーションを介して Akt 経路を活性化し、女性の生殖系列幹細胞の増殖を促進する [印刷に先駆けてオンラインで公開、2022 年 6 月 3 日]。Stem Cell Rev Rep. 2022;10.1007/s12015-022-10394-0。doi:10.1007/s12015-022-10394-0(IF:5.739)
[13] Ru M、Wang W、Zhai Z、et al。ニコチンアミドモノヌクレオチドの補給は、老化マウスとD-ガラクトース誘発老化細胞の腸機能を保護する。Food Funct。2022;13(14):7507-7519。2022年7月18日発行。doi:10.1039/d2fo00525e(IF:5.396)
[14] Ye M、Gao R、Chen S、et al。MEG3のダウンレギュレーションとEZH2のアップレギュレーションが協力して神経芽腫の進行を促進する。J Cell Mol Med。2022;26(8):2377-2391。doi:10.1111/jcmm.17258(IF:5.310)
[15] Wang S, Wang X, Shao Y, et al. 3-(フェニルエチニル)-1,1'-ビフェニル-2-カルボキシレート誘導体の合成と評価、新規HIF-1阻害剤としての研究。Bioorg Chem. 2021;116:105298. doi:10.1016/j.bioorg.2021.105298(IF:5.275)
[16] Guan X、Meng X、Zhu K、et al。MYSM1はRSK3-リン酸化BAD経路を介してアポトーシスを誘導し、TNBC細胞をシスプラチンに対して感受性にする。Cell Death Discov。2022;8(1):84。2022年2月26日発行。doi:10.1038/s41420-022-00881-1(IF:5.241)
[17] Liu H、Wang LL、Xu QH、et al。UHRF1は妊娠初期における栄養芽層の浸潤と脱落膜マクロファージの分化の両方を形作る。FASEB J. 2022;36(4):e22247. doi:10.1096/fj.202101647RR(IF:5.192)
[18] Chen M、Wang J、Lin L、et al. Penicillium brevicompactum における Ca<sup>2+</sup>/活性酸素種による代謝の相乗的制御はミコフェノール酸の産生を改善し、Ca<sup>2+</sup> チャネルの調査を改善する [ACS Synth Biol. 2022 年 4 月 15 日;11(4):1705 に訂正が掲載されました]。ACS Synth Biol. 2022;11(1):273-285. doi:10.1021/acssynbio.1c00413(IF:5.110)
[19] Zhao X、Li X、Yang W、Peng J、Huang J、Mi S.高リスクヒトパピローマウイルスの自動迅速診断のための統合マイクロ流体検出システム。アナリスト。2021;146(16):5102-5114. doi:10.1039/d1an00623a(IF:4.616)
[20] Chen H、Jiang Y、Liu R、et al。クルクミン誘導体C66はJNKを介した炎症の阻害を介して膵臓癌の進行を抑制する。分子。2022;27(10):3076。2022年5月11日発行。doi:10.3390 / moleculars27103076(IF:4.412)
[21] Su W、Qiu J、Mei Y、Zhang XE、He Y、Li F。許容細胞の迅速なスクリーニングによるウイルス分離のためのマイクロ流体細胞チップ[印刷に先駆けてオンラインで公開、2022年5月2日]。Virol Sin。2022;S1995-820X(22)00075-X。doi:10.1016/j.virs.2022.04.011(IF:4.327)
[22] Ding J、Mei S、Cheng W、Ni Z、Yu C。クルクミンはHIFシグナル経路を介してマウスの子宮内膜症を治療する。Am J Transl Res. 2022;14(4):2184-2198。2022年4月15日発行。(IF:4.060)
[23] Zhang XY、Shen HH、Qin XY、et al。IL-27はSTAT3-ESR/PGR制御軸を介して脱落膜化を促進する。J Reprod Immunol. 2022;151:103623. doi:10.1016/j.jri.2022.103623(IF:4.054)
[24] Zhu Q、Muyayalo KP、Xu QH、Wang J、Wang H、Liao AH。Prunella vulgarisはTh1/Th17免疫応答を阻害することで実験的自己免疫甲状腺炎ラットの妊娠転帰を改善できる。J Reprod Immunol. 2022;149:103469. doi:10.1016/j.jri.2021.103469(IF:4.054)
[25] Yang Q、Guo K、Zhou X、et al。細菌感染後の三角帆貝Hyriopsis cumingiiの組織病理学、抗酸化応答、トランスクリプトームおよび遺伝子発現解析。Dev Comp Immunol。2021;124:104175。doi:10.1016 / j.dci.2021.104175(IF:3.636)
[26] Liu Y、Wan HH、Tian DM、et al。遺伝子治療のためのヒスチジンとアルギニンの比率の調節による高効率細胞透過性ペプチドの開発と特性評価。Materials(Basel)。2021;14(16):4674。2021年8月19日発行。doi:10.3390 / ma14164674(IF:3.623)
[27] Fu Y、Wang X、Zhang L、Ren Y、Hao L。移植炎症因子1は糖尿病性腎疾患においてNF-κB経路を介して炎症と酸化ストレスを増強する。Biochem Biophys Res Commun。2022;614:63-69。doi:10.1016/j.bbrc.2022.04.089(IF:3.575)
[28] Zhao Y、Wang XQ。VvMYB1はVvTORプロモーターを制御することでVvTOR遺伝子発現に影響を与え、グルコース蓄積に関与する可能性がある。J Plant Physiol. 2022;272:153668. doi:10.1016/j.jplph.2022.153668(IF:3.549)
[29] Cheng Y, Zheng L, Yang C, Zhang W, Wang H. プロポフォールはlncRNA GAS5をアップレギュレーションすることで神経膠腫細胞の増殖と移動を阻害する。Toxicol In Vitro. 2022;80:105321. doi:10.1016/j.tiv.2022.105321(IF:3.500)
[30] Yang B、Wang F、Zheng G。膜貫通タンパク質TMEM119はWnt/β-カテニン経路を活性化することで乳がん細胞の幹細胞性を促進する。バイオエンジニアリング。2021;12(1):4856-4867。doi:10.1080/21655979.2021.1960464(IF:3.269)
[31] Yang X、Liu X、Song F、et al。野生の地上リス(<em>Spermophilus dauricus</em>)の大腸におけるGPR41とGPR43の季節的発現。Eur J Histochem。2022;66(1):3351。2022年1月21日発行。doi:10.4081/ejh.2022.3351(IF:3.188)
[32] Tang Z、Yuan X、Bai Y、et al。マスクラット(<em>Ondatra zibethicus</em>)の精巣におけるPACAP、VPAC1、VPAC2、PAC1の発現と精巣活動の季節的変化。Eur J Histochem。2022;66(2):3398。2022年5月2日発行。doi:10.4081/ejh.2022.3398(IF:3.188)
[33] Zhu P, Chen Y, Wu F, Meng M, Ji K. Pinus massoniana LambにおけるMEP経路酵素コード遺伝子の発現とプロモーター解析。PeerJ. 2022;10:e13266。2022年4月12日発行。doi:10.7717/peerj.13266(IF:2.984)
[34] Zhai L、Chen W、Cui B、Yu B、Wang Y、Liu H。過剰発現したバーシカンは胃癌における細胞増殖、遊走および浸潤を促進する。Tissue Cell。2021;73:101611。doi:10.1016/j.tice.2021.101611(IF:2.466)
[35] Jiang J、Cai M. カルダモニンは軟骨細胞におけるNrf2/NQO-1シグナル伝達経路の活性化を介してNLRP3インフラマソームを阻害することによりIL-1β誘発性障害を阻害した。J Microbiol Biotechnol. 2021;31(6):794-802. doi:10.4014/jmb.2103.03057(IF:2.351)
支払いとセキュリティ
お支払い情報は安全に処理されます。 クレジットカードの詳細を保存したり、クレジットカード情報にアクセスすることはありません
問い合わせ
あなたも好きかもしれません
よくある質問
この製品は研究目的のみに使用され、人間や動物の治療や診断に使用することを意図したものではありません。製品とコンテンツは、
特定のアプリケーションでは、追加のサードパーティの知的財産権が必要になる場合があります。