



説明
Hieff NGS™ DNA 選択ビーズは、SPRI (固相逆固定化) 原理に基づいて調製されており、次世代シーケンシング (NGS) ライブラリの調製中の DNA 精製およびサイズ選択に適用できます。Hieff NGS™ DNA 選択ビーズは、さまざまな DNA および RNA ライブラリ調製キットと互換性があり、AMPure ビーズの優れた代替品です。
コンポーネント
| コンポーネント番号 | 名前 | 12601ES08 | 12601ES56 | 12601ES75 |
| 12601 | Hieff NGS™ DNA選択ビーズ | 5mL | 60mL | 450mL |
仕様
| 製品ライン | DNA洗浄および選択ビーズ |
| 出発物質 | DNA |
| 互換性 | DNA |
| 絶縁技術 | 磁気ビーズ |
| 最終製品タイプ | DNA |
| 用途(アプリケーション) | DNAセランアップ、DNAサイズ選択 |
配送と保管
ビーズは保冷剤と一緒に発送され、2°C〜8°Cで1年間保管できます。
説明書
- 1. 準備
使用前に選択ビーズを室温で少なくとも 30 分間平衡化します。
- 2. DNAサイズの選択
サイズ選択の操作フローを図 1 に示します。プロトコルは次のとおりです。
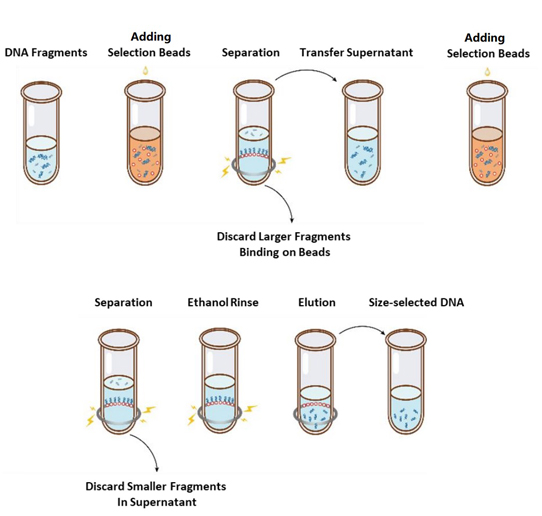
図1. DNAサイズ選択のフローチャート
2.1 使用する前に毎回、ボルテックスまたはピペッティングでビーズをよく混ぜます。
2.2 サンプルに最初の選択ビーズを加えます(表 1 を参照)。ボルテックスまたはピペッティングで少なくとも 10 回上下させてよく混ぜます。
2.3 室温で5分間インキュベートします。
2.4 チューブを軽くスピンダウンし、磁気スタンドに置きます。溶液が透明になったら(約 5 分)、上清を新しい PCR チューブに移します。
2.5 表 1 に従って、ステップ 2.4 のサンプルに 2 回目の選択ビーズを追加します。少なくとも 10 回ボルテックスまたはピペッティングで完全に混合します。
2.6 室温で5分間インキュベートします。
2.7 チューブを軽くスピンダウンし、磁気スタンドに置きます。溶液が透明になったら(約 5 分)、上清を吸引して捨てます。
2.8 チューブを磁気スタンドに置いたまま、ビーズを乱さないように、新しく調製した 80% エタノール 200 μL を加え、室温で 30 秒間インキュベートします。エタノールを吸引して捨てます。
2.9 手順 2.8 をもう一度繰り返して、合計 2 回洗浄します。
2.10 10 µL ピペットチップで残留エタノールを除去します。チューブを磁気スタンドに置いたまま、蓋を開けた状態で選択ビーズをひび割れが現れるまで(約 5 分)空気乾燥させます。
注意: 選択ビーズを乾燥させすぎないでください。DNA ターゲットの回収率が低下する可能性があります。
2.11 チューブを磁気スタンドから取り外し、適量の ddH2O (≥20 µL) を加え、少なくとも 10 回ボルテックスまたはピペッティングでよく混ぜます。
2.12 室温で5分間インキュベートする。
チューブを軽くスピンダウンし、磁気スタンドに置きます。溶液が透明になったら(約 5 分)、上清 20 μL を新しいチューブに移します。
- 3.DNAサイズ選択の推奨条件
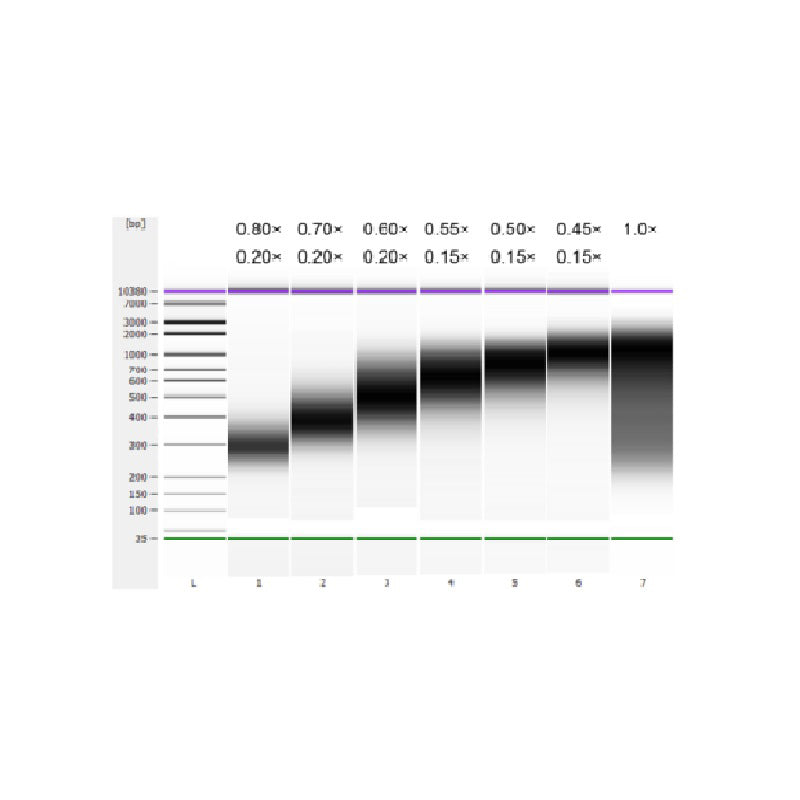
子牛胸腺 DNA を超音波処理により断片化して 100 ~ 1,000 bp の断片を調製し、表 1 に従って 2 回のサイズ選択を実行しました。結果は Agilent 2100 Bioanalyzer を使用して分析しました (図 2)。
表1. DNAサイズ選択の推奨条件
| DNA断片の長さ | 250-350bp | 320-420bp | 450-550bp | 550-700bp | 700-900bp | 800~1,000年前 |
| ビーズとDNAの比率(第1ラウンド) | 0.80× | 0.70× | 0.60× | 0.55× | 0.50× | 0.45× |
| 第2ラウンドのビーズとDNAの比率 | 0.20× | 0.20× | 0.20× | 0.15倍 | 0.15倍 | 0.15倍 |
注: 表中の「×」はサンプル DNA の容量を示します。たとえば、ライブラリの挿入長が 250 bp でサンプル DNA の容量が 100 μL の場合、1 回目の分類で使用する磁気ビーズの容量は 0.80×100 μL=80 μL、2 回目の分類で使用する磁気ビーズの容量は 0.20×100 μL=20 μL です。
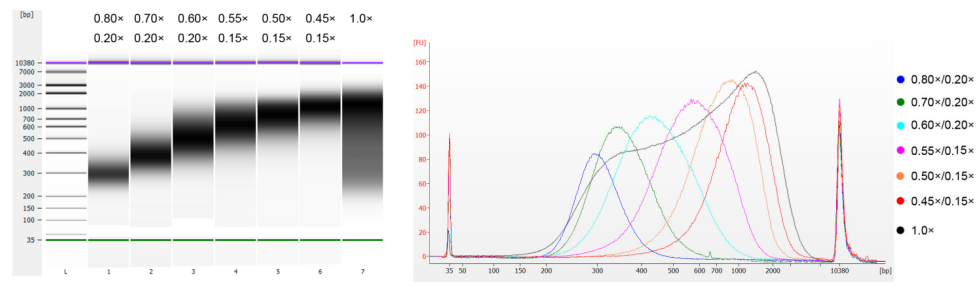
図2. Agilent 2100高感度DNAチップの電気泳動図
注:
1. 安全と健康のため、操作時には白衣と使い捨て手袋を着用してください。

「Candidatus Nitrosopumilus koreensis AR1由来のファミリー1カスポザーゼによる配列特異的統合。Nucleic Acids Res. 2021;49(17):9938-9952. doi:10.1093/nar/gkab725」より引用

「最近のウォルバキア感染により野生のラオデルファクス・ストリアテルス個体群の微生物群集が変化。マイクロバイオーム。2020;8(1):104。2020年7月2日発行。doi:10.1186/s40168-020-00878-x」より引用
[1] Wang X、Yuan Q、Zhang W、et al. Candidatus Nitrosopumilus koreensis AR1由来のファミリー1カスポザーゼによる配列特異的統合。Nucleic Acids Res. 2021;49(17):9938-9952. doi:10.1093/nar/gkab725(IF:16.971)
[2] Duan XZ、Sun JT、Wang LT、et al。最近のWolbachia感染は野生のLaodelphax striatellus個体群の微生物群集を変化させる。Microbiome。2020;8(1):104。2020年7月2日発行。doi:10.1186/s40168-020-00878-x(IF:11.607)
[3] Song B、Almatrafi E、Sang F、et al。バイオ炭と羊糞堆肥によるフェントン処理堆積物の管理:細菌群集の進化特性への影響。J Environ Manage。2022;316:115218。doi:10.1016/j.jenvman.2022.115218(IF:6.789)
[4] Huang C、Mei Q、Lou L、et al。腸内細菌叢とTh17/Treg細胞バランスの調整による糞便細菌叢移植に対する潰瘍性大腸炎の反応。細胞。2022;11(11):1851。2022年6月5日発行。doi:10.3390/cells11111851(IF:6.600)
[5] Ghosh S、Yang X、Wang L、Zhang C、Zhao L。活性期プレバイオティクス摂取は腸内細菌叢を変化させ、高脂肪食を与えられたマウスの体重非依存的な脂肪肝および血清コレステロールの緩和を誘導する。Comput Struct Biotechnol J. 2020;19:448-458。2020年12月24日発行。doi:10.1016/j.csbj.2020.12.011(IF:6.018)
[6] Gao X、Yu B、Yu J、et al。子豚の食事性炭水化物消化の発達プロファイリング。Front Microbiol。2022;13:896660。2022年4月29日発行。doi:10.3389/fmicb.2022.896660(IF:5.640)
[7] Li P、Zhang Y、Yan F、Zhou X。臨床Klebsiella oxytocaから分離されたバクテリオファージvB_Kox_ZX8の特性とマウス菌血症に対する治療効果。Front Microbiol。2021;12:763136。2021年12月3日発行。doi:10.3389/fmicb.2021.763136(IF:5.640)
[8] Lin Z、Luo P、Huang D、Wu Y、Li F、Liu H。サッカロミセス・セレビシエモデルにおけるアクリルアミドの毒性解析のためのマルチオミクスベースの戦略。Chem Biol Interact。2021;349:109682。doi:10.1016 / j.cbi.2021.109682(IF:5.194)
[9] Sun X、Lv W、Wang Y 他。 Mrgprb2 遺伝子は、ドクダミ注射によって誘発されるアナフィラキシー様反応において役割を果たします。 J エスノファーマコル。 2022;289:115053。 doi:10.1016/j.jep.2022.115053(IF:4.360)
[10] Ma H、Lai B、Zan C、Di X、Zhu X、Wang K. GLO1はPERタイプの拡張スペクトルβ-ラクタマーゼを誘導することにより大腸菌の薬剤耐性に寄与する。感染薬剤耐性。2022;15:1573-1586。2022年4月5日発行。doi:10.2147/IDR.S358578(IF:4.003)
[11] Zhong Y、Zhao W、Tang Z、et al。アカザリガニProcambarus clarkiiの卵巣の異なる発達段階の比較トランスクリプトーム解析。BMC Genomics。2021;22(1):199。2021年3月21日発行。doi:10.1186/s12864-021-07537-x(IF:3.969)
[12] Lian C、Yang H、Lan J、et al. 葉緑体ゲノムの比較分析により、薬用植物Isodon rubescensの系統関係と種内変異が明らかになった。PLoS One. 2022;17(4):e0266546. 2022年4月6日発行。doi:10.1371/journal.pone.0266546(IF:3.240)
[13] Diao G、Huang J、Zheng X、et al。プロスタグランジンE2は樹状細胞の移動を制御する上で二重の役割を果たしている。Int J Mol Med。2021;47(1):207-218。doi:10.3892/ijmm.2020.4801(IF:3.098)
[14] Bing XL、Zhao DS、Peng CW、Huang HJ、Hong XY。イネウンカ(半翅目:ウンカ科)個体群における細菌および真菌群集の類似性と空間的変動。Insect Sci. 2020;27(5):947-963. doi:10.1111/1744-7917.12782(IF:2.791)
[15] Li X、Zhou S、Zhang J、Zhou Z、Xiong Q。クロコウチュウバエ(Hermetia illucens)幼虫の腸内細菌群集の方向性変化。動物(バーゼル)。2021;11(12):3475。2021年12月6日発行。doi:10.3390/ani11123475(IF:2.752)
[16] Yang J、Peng Y、Kong W. 桑の潜在性ウイルス1の同定と完全なゲノム配列。Arch Virol. 2022;167(2):687-690. doi:10.1007/s00705-021-05350-1(IF:2.574)
[17] Chang Y、Xia X、Sui L、et al。昆虫病原菌の内生コロニー形成は、内生細菌群集を変化させることで植物の病害抵抗性を高める。J Basic Microbiol. 2021;61(12):1098-1112. doi:10.1002/jobm.202100494(IF:2.281)
[18] Ding CY、Ma YM、Li B、et al. Myzus persicae(半翅目:アブラムシ科)におけるトランスアネトールに対する応答における発現差遺伝子の同定と機能解析。J Insect Sci. 2022;22(1):3. doi:10.1093/jisesa/ieab094(IF:1.857)
支払いとセキュリティ
お支払い情報は安全に処理されます。 クレジットカードの詳細を保存したり、クレジットカード情報にアクセスすることはありません
問い合わせ
あなたも好きかもしれません
よくある質問
この製品は研究目的のみに使用され、人間や動物の治療や診断に使用することを意図したものではありません。製品とコンテンツは、
特定のアプリケーションでは、追加のサードパーティの知的財産権が必要になる場合があります。