

説明
GMyc-PCR マイコプラズマ検出キットは、主にPCR法を使用して、さまざまな生物学的材料(細胞培養、実験動物の分泌物、動物血清など)のマイコプラズマ感染を検出します。感度、特異性、迅速性などの利点を兼ね備えており、細胞培養上清で直接検出できます。この製品は、PCR法で培養細胞などの生物学的材料内のマイコプラズマを検出します。使用するプライマーは、マイコプラズマの16S-23S rRNA配列の保存領域に従って設計されており、マイコプラズマDNAのみを特異的に増幅し、検出感度と特異性が高くなっています。PCR増幅と電気泳動分析には数時間しかかからず、操作は便利で簡単です。
細胞培養は生命科学研究において一般的な実験です。他の一般的な実験方法とは異なり、細胞培養は動的な連続プロセスであり、細胞は操作エラーや汚染物質に反応して、異常な細胞状態や培地の外観を示すことがよくあります。マイコプラズマに汚染されている場合、細胞形態に明らかな変化はなく、見落とされがちです。汚染が非常に深刻になるまで発見されないことがよくあります。汚染された細胞膜には数百のマイコプラズマが存在する可能性があり、これらのマイコプラズマは栄養素を奪い合い、有毒な代謝物を放出し、実験結果に重大な影響を及ぼします。
研究によると、少なくとも 20 種類のマイコプラズマが細胞を汚染する可能性があり、その中で最も一般的なものは、口腔マイコプラズマ (M. orale)、マイコプラズマ アルギニン ( M. arginini )、マイコプラズマ ヒオリニス ( M. hyorhinis)、マイコプラズマ ファーメンタム ( M. fermentans )、マイコプラズマ ホミニス ( M. hominis )、マイコプラズマ サリバリウス (M. salivarium )、マイコプラズマ プルモナリー ( M. pulmonis )、マイコプラズマ ペア ( M. pirum ) です。培養細胞のマイコプラズマ汚染率は 4% から 92% の範囲です。汚染源としては、作業環境、作業者自身(一部のマイコプラズマは人体の常在菌です)、培養培地、血清、細胞の交差汚染、実験装置、細胞を調製した元の組織または臓器の使用済み汚染などが挙げられます。
細胞培養中に生じる問題の根本原因を特定することは困難で時間のかかる作業であり、突然の変化を疑う必要があり、適切な検査方法とマイコプラズマ汚染の定期的な検査が必要です。マイコプラズマの検出には、直接培養、DNA蛍光染色、ELISA、PCR法など、さまざまな方法があります。
イェーセン マイコプラズマ汚染に対する包括的なソリューションを提供します。関連製品をご覧ください: マイコプラズマの概念と汚染の影響
特徴
- 使用したプライマーは、マイコプラズマ16S-23S rRNA配列の保存領域に従って設計された。
- 増幅されたマイコプラズマDNAのみが特異的に
- 優れた感度と特異性
- 20種以上のマイコプラズマが検出されました
応用
- マイコプラズマ検出
コンポーネント
| コンポーネント番号 | 名前 | 40601ES10 (10アッセイ) | 40601ES20 (20アッセイ) |
| 40601-A | GMyc-1st PCR ミックス | 250µL | 2×250µL |
| 40601-B | GMyc-2nd PCR ミックス | 250µL | 2×250µL |
| 40601-C | 陽性コントロールテンプレート N | 20µL | 20µL |
【注意事項】 1.長期間使用しない場合は、-85~-65℃で冷凍保存できます。
2. PCR 反応は非常に敏感です。偽陽性を防ぐために、サンプルを追加するときに最後に陽性コントロールを追加します。
ストレージ
この製品は-25〜-15℃で18ヶ月間保存できます。長期間使用しない場合は、光を避けて保管してください。
数字
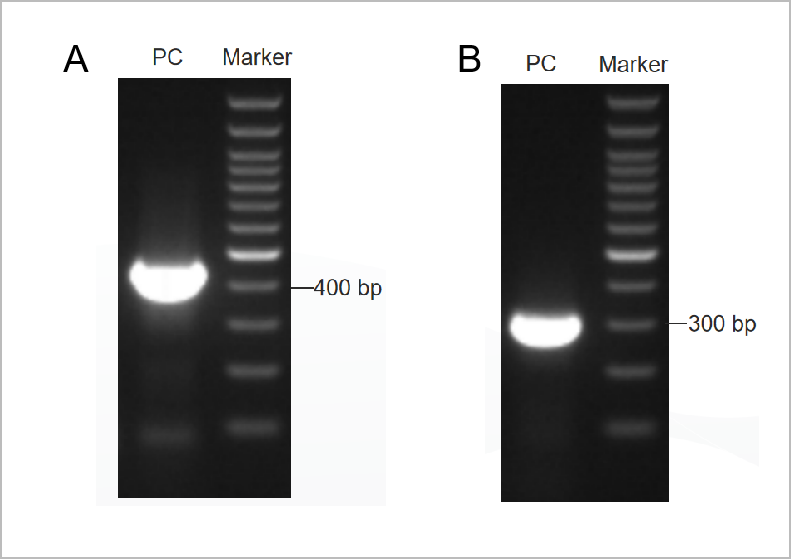
図1. PCRマイコプラズマ検査キットの電気泳動図の結果。
1 回目の電気泳動図を図 1A に示し、2 回目の電気泳動図を図 1B に示します。(M: 1kb マーカー、1-2: 陽性コントロール、3: 陰性コントロール)
[1] Rao XS、Cong XX、Gao XK、et al. AMPKを介したリン酸化はTBC1D17の自己阻害を促進し、Rab5依存性のグルコース取り込みを促進する。Cell Death Differ. 2021;28(12):3214-3234. doi:10.1038/s41418-021-00809-9(IF:15.828)
[2] Guo F, Li L, Li J, et al. マウス初期胚および胚性幹細胞の単一細胞マルチオミクスシークエンシング。Cell Res. 2017;27(8):967-988. doi:10.1038/cr.2017.82(IF:15.606)
[3] Hao Y, He B, Wu L, et al. p85βの核内転座はPIK3CAヘリカルドメイン変異癌の腫瘍形成を促進する。Nat Commun. 2022;13(1):1974. 2022年4月13日発行。doi:10.1038/s41467-022-29585-x(IF:14.919)
[4] Shu X、Liu M、Lu Z、et al.ゲノムワイドマッピングにより、デオキシウリジンがヒトセントロメアDNAに豊富に含まれていることが明らかになりました。Nat Chem Biol. 2018;14(7):680-687. doi:10.1038/s41589-018-0065-9(IF:13.843)
[5] Li X、Xiong X、Wang K、et al. トランスクリプトーム全体のマッピングにより、可逆的かつ動的なN(1)-メチルアデノシンメチロームが明らかになった。Nat Chem Biol. 2016;12(5):311-316. doi:10.1038/nchembio.2040(IF:12.709)
[6] Sun L、Yang X、Huang X、et al。脂肪酸の2-ヒドロキシル化はYAP転写軸を介して大腸腫瘍形成および転移を抑制する。Cancer Res。2021;81(2):289-302。doi:10.1158/0008-5472.CAN-20-1517(IF:12.701)
[7] Sun Z、Zhang Z、Wang QQ、Liu JL。CTPS1とATRの複合不活性化はMYC過剰発現癌細胞に対して合成的に致死的である。Cancer Res. 2022;82(6):1013-1024. doi:10.1158/0008-5472.CAN-21-1707(IF:12.701)
[8] Song J、Zhuang Y、Zhu C、et al. miRNA処理とtRNA擬似ウリジル化におけるヒトPUS10の異なる役割。Nat Chem Biol. 2020;16(2):160-169. doi:10.1038/s41589-019-0420-5(IF:12.154)
[9] He B、Pan H、Zheng F、et al。長鎖非コードRNA LINC00930は、鼻咽頭癌におけるPFKFB3を介した腫瘍解糖および細胞増殖を促進する。J Exp Clin Cancer Res。2022;41(1):77。2022年2月24日発行。doi:10.1186/s13046-022-02282-9(IF:11.161)
[10] Tang B、Liu BH、Liu ZY、Luo MY、Shi XH、Pang DW。コンパクトな両親媒性双性イオンコーティングを施した量子ドット。ACS Appl Mater Interfaces。2022;14(24):28097-28104。doi:10.1021/acsami.2c04438(IF:9.229)
[11] Huang C、Zhang Z、Chen L、et al. SrcのN末端ドメインとC末端ドメイン内のアセチル化はSTAT3を介した腫瘍形成の異なる役割を制御する。Cancer Res. 2018;78(11):2825-2838. doi:10.1158/0008-5472.CAN-17-2314(IF:9.130)
[12] Wu X、Yu M、Zhang Z、et al。DDB2はCDT2のPCNA非依存性分解を介してDNA複製を制御する。Cell Biosci。2021;11(1):34。2021年2月8日発行。doi:10.1186 / s13578-021-00540-5(IF:7.133)
[13] Wang J、Zhang Y、Liu X、Liu H。腫瘍抵抗性サブポピュレーションへの到達可能性に基づく適応療法の最適化。Cancers(Basel)。2021;13(21):5262。2021年10月20日発行。doi:10.3390 / cancers13215262(IF:6.639)
[14] Feng W, Liu R, Xie X, et al. α-チューブリンのSUMO化は微小管のダイナミクスを制御する新しい修飾である。J Mol Cell Biol. 2021;13(2):91-103. doi:10.1093/jmcb/mjaa076(IF:6.216)
[15] Yu M、Hu X、Yan J、Wang Y、Lu F、Chang J。RIOK2阻害剤NSC139021は、Skp2を介した細胞周期停止とアポトーシスを誘導することにより、神経膠芽腫に対して抗腫瘍効果を発揮します。バイオメディシンズ。2021;9(9):1244。2021年9月17日発行。doi:10.3390/biomedicines9091244(IF:6.081)
[16] Ren S、Cai Y、Hu S、et al. ベルベリンはc-myc/CD47軸を調節することでびまん性大細胞型B細胞リンパ腫に抗腫瘍活性を発揮する。Biochem Pharmacol. 2021;188:114576. doi:10.1016/j.bcp.2021.114576(IF:5.858)
[17] Wen F, Sun X, Sun C, et al. TAGLNは前立腺癌細胞においてTRAF6を介したプロテアソーム分解によってダウンレギュレーションされる。Mol Cancer Res. 2021;19(7):1113-1122. doi:10.1158/1541-7786.MCR-20-0513(IF:5.852)
[18] Tang B、Sun EZ、Zhang ZL、et al. スフィンゴミエリン隔離コレステロールドメインは、インフルエンザウイルスの侵入時にクラスリン被覆ピットを収縮させるためにフォルミン結合タンパク質17をリクルートする。J Virol. 2022;96(5):e0181321. doi:10.1128/JVI.01813-21(IF:5.103)
[19] Hu J、Ren W、Qiu W、et al。JAK2変異を有する強直性脊椎炎患者からの誘導多能性幹細胞株(XDCMHi001-A)の生成。Stem Cell Res。2020;45:101788。doi:10.1016 / j.scr.2020.101788(IF:4.495)
[20] Xiao S、Yao X、Ye J、Tian X、Yin Z、Zhou L。エピジェネティック修飾は胃癌におけるプロリン合成酵素PYCR1の異常発現を促進する[印刷に先駆けてオンライン公開、2022年5月30日]。Biochim Biophys Acta Gene Regul Mech。2022;1865(6):194829。doi:10.1016/j.bbagrm.2022.194829(IF:4.490)
[21] Wang J、Zhang Y、Liu X、Liu H。完全な情報がない場合、固定周期治療は腫瘍システムに効果的ですか?Cancer Manag Res。2021;13:8915-8928。2021年11月30日発行。doi:10.2147 / CMAR.S339787(IF:3.989)
[22] Yang X、Ren S、Rehman ZU、et al. 牛(Bos taurus)とヤギ(Capra hircus)のTANK結合キナーゼ1(TBK1)の分子特性、発現、機能同定。Dev Comp Immunol. 2022;133:104444. doi:10.1016/j.dci.2022.104444(IF:3.636)
[23] Zheng D, Chang X, Liu Y, et al. 2-メトキシ-5((3,4,5-トリメトシフェニル)セレニニル)フェノールは、乳がん細胞株におけるMDM2のダウンレギュレーションを介してEGF誘導性細胞遊走および浸潤を逆転させる。Cancer Biol Ther. 2019;20(4):513-523. doi:10.1080/15384047.2018.1537578(IF:3.373)
[24] Xu F, Zhang S, Liu Z, et al. TEX9とeIF3bは機能的に相乗効果を発揮し、食道扁平上皮癌の進行を促進する。BMC Cancer. 2019;19(1):875. 2019年9月3日発行。doi:10.1186/s12885-019-6071-9(IF:2.933)
[25] Pan H、Sun L、Wang W、et al。血清長鎖非コードRNA LOC553103は、一般的なタイプのヒト癌の非特異的診断および予後バイオマーカーとして機能します。Clin Chim Acta。2020;508:69-76。doi:10.1016 / j.cca.2020.05.017(IF:2.615)
[26] Li N、Lin SM、Li Y、Sun J、Zhang L、Chen M。軟骨無形成症を引き起こすFGFR3遺伝子のヘテロ接合性G380R変異を持つ胎児由来の誘導多能性幹細胞株(GZHMCi004-A)。Stem Cell Res。2021;53:102322。doi:10.1016 / j.scr.2021.102322(IF:2.020)
[27] Luo Q, Wei C, Long Y, et al. iCRISPR/Cas9システムによるELTD1ノックアウトヒト胚性幹細胞株の生成。Stem Cell Res. 2021;53:102350. doi:10.1016/j.scr.2021.102350(IF:2.020)
[28] Liu YQ、Ling TW、Wang HY、Yang YH、Song WJ、Wang TC。DGKG変異を有するてんかん患者からの統合フリー誘導多能性幹細胞株(LZUSHI001-A)の生成。Stem Cell Res。2022;61:102768。doi:10.1016 / j.scr.2022.102768(IF:2.020)
[29] Chen M、Lin SM、Li N、Li Y、Li Y、Zhang L。RUNX2遺伝子のエクソン3ヘテロ欠失を伴う胎児由来の誘導多能性幹細胞株(GZHMCi003-A)は、鎖骨頭蓋異形成症を引き起こします。Stem Cell Res. 2021;51:102166. doi:10.1016/j.scr.2021.102166(IF:2.020)
[30] Xu Y、Wang X、Qiu T、et al. DNM1遺伝子にヘテロ接合性c.545C>A変異を有する発達性てんかん性脳症31型患者由来の誘導多能性幹細胞株(FDCHI007-A)の生成。Stem Cell Res. 2022;60:102709. doi:10.1016/j.scr.2022.102709(IF:2.020)
[31] Fan T、He J、Wang Y、Yu J、Sun W。中枢性早発思春期の7歳女児からの誘導多能性幹細胞株(FDCHi006-A)の生成。Stem Cell Res。2021;56:102542。doi:10.1016 / j.scr.2021.102542(IF:2.020)
[32] Gong X、Zheng Z、Yang T、Zheng H、Xiao X、Jia N. ARX遺伝子のヘミ接合性R330L変異を有する1型早期乳児てんかん性脳症(EIEE1)患者からの同質遺伝子修正iPSCライン(OGHFUi001-A-1)の生成。Stem Cell Res. 2022;60:102693. doi:10.1016/j.scr.2022.102693(IF:2.020)
[33] Jia N、Gong X、Chen J、et al。ARX変異を伴う1型早期乳児てんかん性脳症からの誘導多能性幹細胞株(OGHFUi001-A)の生成。Stem Cell Res。2021;53:102367。doi:10.1016 / j.scr.2021.102367(IF:2.020)
[34] Zhu W、Zhou Y、Wang Q、et al. PSEN1遺伝子にE363Q変異を有する患者の皮膚線維芽細胞からのヒト誘導多能性幹細胞(iPSC)株の生成。Stem Cell Res. 2022;61:102769. doi:10.1016/j.scr.2022.102769(IF:2.020)
[35] Luo F, Long K, Li X, et al. LRRC8Aの欠損は3T3-L1細胞における低酸素誘発壊死を軽減する。Biosci Biotechnol Biochem. 2020;84(6):1139-1145. doi:10.1080/09168451.2020.1730689(IF:1.516)
支払いとセキュリティ
お支払い情報は安全に処理されます。 クレジットカードの詳細を保存したり、クレジットカード情報にアクセスすることはありません
問い合わせ
あなたも好きかもしれません
よくある質問
この製品は研究目的のみに使用され、人間や動物の治療や診断に使用することを意図したものではありません。製品とコンテンツは、
特定のアプリケーションでは、追加のサードパーティの知的財産権が必要になる場合があります。