





Beschrijving
Hieff NGS™ DNA Selection Beads worden bereid op basis van het SPRI (Solid Phase Reverse Immobilization) principe en zijn toepasbaar voor DNA-zuivering en grootteselectie tijdens de voorbereiding van next generation sequencing (NGS) bibliotheken. Hieff NGS™ DNA Selection Beads zijn compatibel met verschillende DNA- en RNA-bibliotheekvoorbereidingskits en vormen een goed alternatief voor AMPure kralen.
Componenten
| Componenten nr. | Naam | 12601ES08 | 12601ES56 | 12601ES75 |
| 12601 | Hieff NGS™ DNA-selectiekralen | 5 ml | 60 ml | 450 ml |
Specificaties
| Productlijn | DNA-schoonmaak en selectie kralen |
| Uitgangsmateriaal | DNA |
| Verenigbaarheid | DNA |
| Isolatietechnologie | Magnetische kraal |
| Eindproducttype | DNA |
| Voor gebruik met (toepassing) | DNA cel omhoog, DNA grootte selectie |
Verzending en opslag
De kralen worden geleverd met koelelementen en kunnen een jaar lang bewaard worden bij een temperatuur van 2°C-8°C.
Instructies
- 1. Voorbereiding
Laat de geselecteerde kralen minimaal 30 minuten op kamertemperatuur komen voordat u ze gebruikt.
- 2. DNA-grootteselectie
De operationele stroom van de grootteselectie wordt weergegeven in Figuur 1 en het protocol is als volgt.
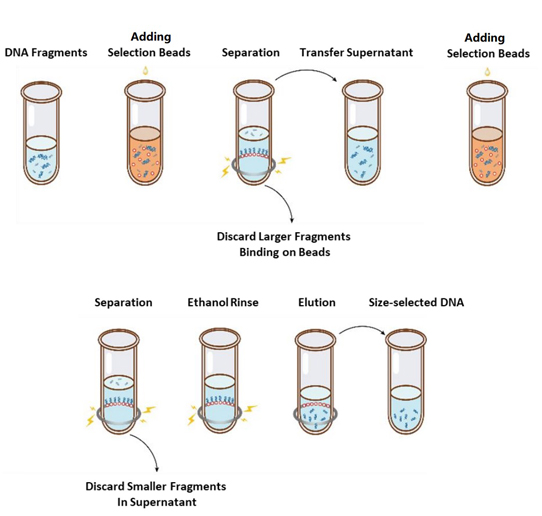
Figuur 1. Het stroomdiagram van DNA-grootteselectie
2.1 Meng de kralen grondig door ze voor gebruik te vortexen of op en neer te pipetteren.
2.2 Voeg de eerste ronde selectieparels toe aan het monster (zie Tabel 1). Meng grondig door te vortexen of door minstens 10 keer op en neer te pipetteren.
2.3 Laat 5 min. bij kamertemperatuur incuberen.
2.4 Draai de buis kort en plaats deze op een magnetische standaard. Wanneer de oplossing helder is (ongeveer 5 min), breng de supernatant over naar een nieuwe PCR-buis.
2.5 Voeg de tweede ronde selectieparels toe aan het monster uit stap 2.4 volgens Tabel 1. Meng grondig door te vortexen of door minstens 10 keer op en neer te pipetteren.
2.6 Laat 5 min. bij kamertemperatuur incuberen.
2.7 Draai de buis kort en plaats deze op een magnetische standaard. Wanneer de oplossing helder is (ongeveer 5 min), zuig de supernatant op en gooi deze weg.
2.8 Houd de buis in de magnetische standaard en voeg 200 μL vers bereide 80% ethanol toe zonder de kralen te verstoren, incubeer bij kamertemperatuur gedurende 30 sec. Zuig de ethanol op en gooi deze weg.
2.9 Herhaal stap 2.8 eenmaal voor in totaal twee wasbeurten.
2.10 Verwijder restanten ethanol met 10 µL pipetpunten. Houd de buis in de magnetische standaard, laat de selectiekralen aan de lucht drogen met het deksel open totdat er net barsten verschijnen (ongeveer 5 min).
Let op: Droog de selectiekralen niet te lang. Dit kan resulteren in een lager DNA-doelwit.
2.11 Haal de buis uit de magnetische standaard. Voeg een geschikte hoeveelheid ddH2O (≥20 µL) toe en meng grondig door te vortexen of door minstens 10 keer op en neer te pipetteren.
2.12 Laat 5 min. bij kamertemperatuur incuberen.
Draai de buis even omlaag en plaats deze op de magnetische standaard. Wanneer de oplossing helder is (ongeveer 5 minuten), breng dan 20 μL van de supernatant over naar een nieuwe buis.
- 3. Aanbevolen omstandigheden voor DNA-grootteselectie
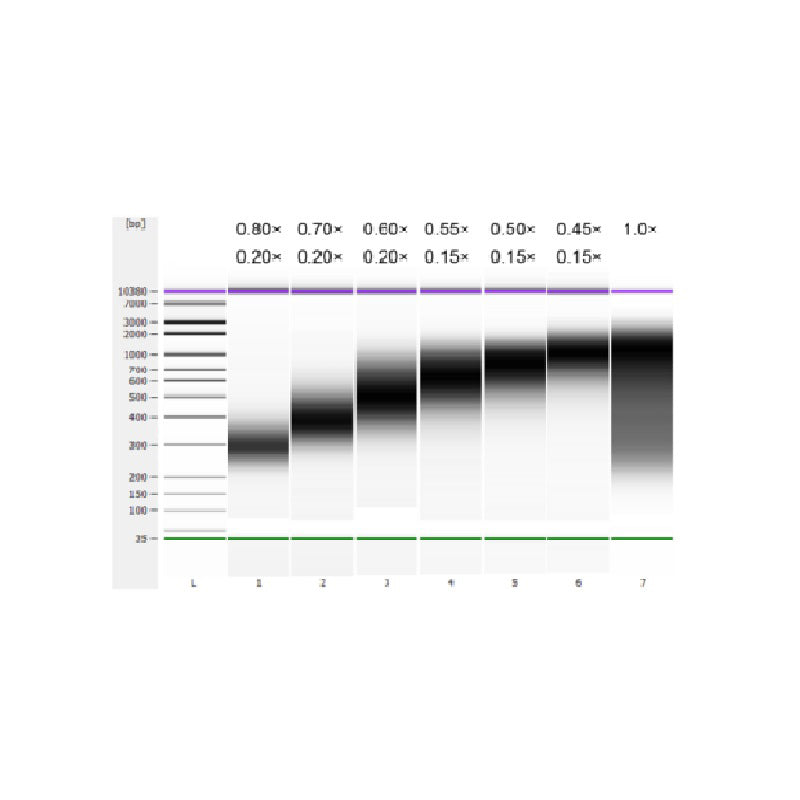
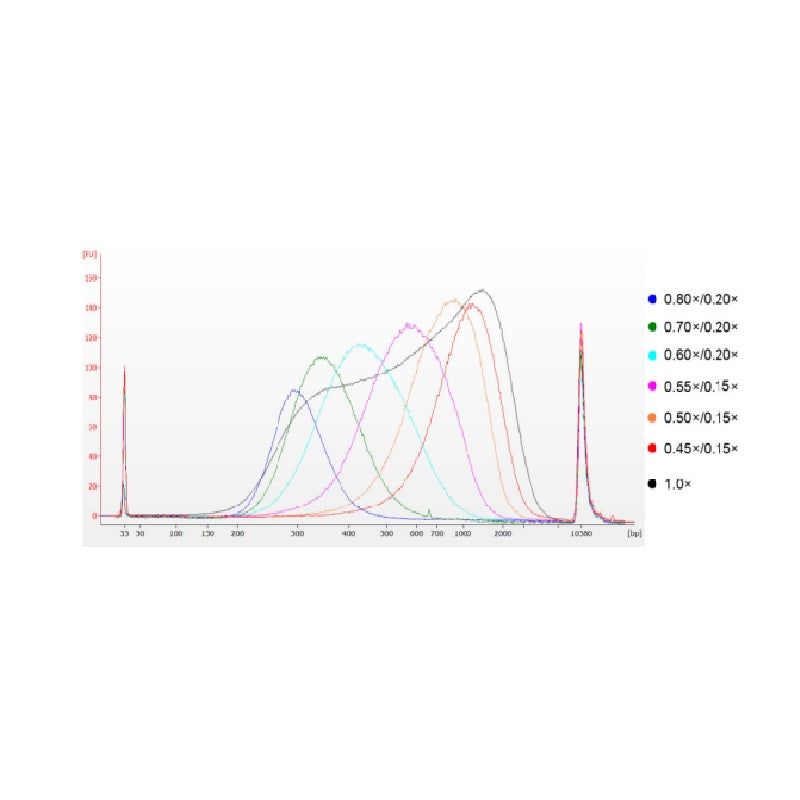
Het kalfsthymus-DNA werd gefragmenteerd door middel van sonificatie om een fragment van 100-1.000 bp te bereiden, en er werden twee ronden van grootteselectie uitgevoerd volgens Tabel 1. De resultaten werden geanalyseerd met behulp van Agilent 2100 Bioanalyzer (Figuur 2).
Tabel 1. Aanbevolen conditie voor DNA-grootteselectie
| Lengte van DNA-fragment | 250-350 basispunten | 320-420 basispunten | 450-550 jaar geleden | 550-700 basispunten | 700-900 basispunten | 800-1.000 basispunten |
| Verhouding kralen: DNA voor de 1e ronde | 0,80× | 0,70× | 0,60× | 0,55× | 0,50× | 0,45× |
| Verhouding kralen: DNA voor de 2e Ronde | 0,20× | 0,20× | 0,20× | 0,15× | 0,15× | 0,15× |
Opmerking: "×" in de tabel geeft het volume van het DNA-monster aan. Als de insertlengte van de bibliotheek bijvoorbeeld 250 bp is en het DNA-monstervolume 100 μL, is het volume van de magnetische kralen die in de eerste sorteerronde worden gebruikt 0,80 × 100 μL = 80 μL; het volume van de magnetische kralen die in de tweede sorteerronde worden gebruikt, is 0,20 × 100 μL = 20 μL.
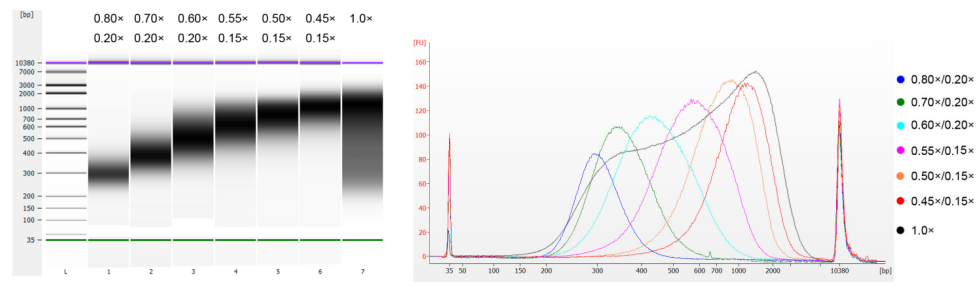
Figuur 2. Agilent 2100 DNA-chip-elektroferogram met hoge gevoeligheid
Opmerkingen:
1. Voor uw eigen veiligheid en gezondheid verzoeken wij u om tijdens de operatie een labjas en wegwerphandschoenen te dragen.

Geciteerd uit "Sequentiespecifieke integratie door de familie 1 casposase van Candidatus Nitrosopumilus koreensis AR1. Nucleïnezuren Res. 2021;49(17):9938-9952. doi:10.1093/nar/gkab725"

Geciteerd uit "Recente infectie door Wolbachia verandert microbiële gemeenschappen in wilde Laodelphax striatellus-populaties. Microbiome. 2020;8(1):104. Gepubliceerd 2020 Jul 2. doi:10.1186/s40168-020-00878-x"
[1] Wang X, Yuan Q, Zhang W, et al. Sequentiespecifieke integratie door de familie 1 casposase van Candidatus Nitrosopumilus koreensis AR1. Nucleïnezuren Res. 2021;49(17):9938-9952. doi:10.1093/nar/gkab725(IF:16.971)
[2] Duan XZ, Sun JT, Wang LT, et al. Recente infectie door Wolbachia verandert microbiële gemeenschappen in wilde Laodelphax striatellus-populaties. Microbiome. 2020;8(1):104. Gepubliceerd 2020 jul. 2. doi:10.1186/s40168-020-00878-x(IF:11.607)
[3] Lied B, Almatrafi E, Sang F, et al.Beheer van Fenton-behandeld sediment met biochar en schapenmestcompost: effecten op de evolutionaire kenmerken van de bacteriële gemeenschap. J Environ Manage. 2022;316:115218. doi:10.1016/j.jenvman.2022.115218(IF:6.789)
[4] Huang C, Mei Q, Lou L, et al. Colitis ulcerosa als reactie op fecale microbiotatransplantatie via modulatie van darmmicrobiota en Th17/Treg-celbalans. Cells. 2022;11(11):1851. Gepubliceerd op 5 juni 2022. doi:10.3390/cells11111851(IF:6.600)
[5] Ghosh S, Yang X, Wang L, Zhang C, Zhao L. Actieve fase prebiotische voeding verandert darmflora, induceert gewichtsonafhankelijke verlichting van leververvetting en serumcholesterol bij muizen die een vetrijk dieet kregen. Comput Struct Biotechnol J. 2020;19:448-458. Gepubliceerd 2020 Dec 24. doi:10.1016/j.csbj.2020.12.011(IF:6.018)
[6] Gao X, Yu B, Yu J, et al. Ontwikkelingsprofiel van de vertering van koolhydraten in het dieet bij biggen. Front Microbiol. 2022;13:896660. Gepubliceerd op 29 april 2022. doi:10.3389/fmicb.2022.896660(IF:5.640)
[7] Li P, Zhang Y, Yan F, Zhou X. Kenmerken van een bacteriofaag, vB_Kox_ZX8, geïsoleerd uit klinische Klebsiella oxytoca en het therapeutische effect ervan op muizenbacteriëmie. Front Microbiol. 2021;12:763136. Gepubliceerd 3 dec. 2021. doi:10.3389/fmicb.2021.763136(IF:5.640)
[8] Lin Z, Luo P, Huang D, Wu Y, Li F, Liu H. Multi-omics-gebaseerde strategie voor toxiciteitsanalyse van acrylamide in het Saccharomyces cerevisiae-model. Chem Biol Interact. 2021;349:109682. doi:10.1016/j.cbi.2021.109682(IF:5.194)
[9] Zon X, Lv W, Wang Y, et al. Het Mrgprb2-gen speelt een rol bij de anafylactoïde reacties veroorzaakt door injectie met Houttuynia cordata. J Ethnopharmacol. 2022;289:115053. doi:10.1016/j.jep.2022.115053(IF:4.360)
[10] Ma H, Lai B, Zan C, Di X, Zhu X, Wang K. GLO1 draagt bij aan de medicijnresistentie van Escherichia coli door het induceren van PER-type van Extended-Spectrum β-Lactamases. Infect Drug Resist. 2022;15:1573-1586. Gepubliceerd 2022 Apr 5. doi:10.2147/IDR.S358578(IF:4.003)
[11] Zhong Y, Zhao W, Tang Z, et al. Vergelijkende transcriptoomanalyse van de verschillende ontwikkelingsstadia van de eierstokken bij de rode moeraskreeft Procambarus clarkii. BMC Genomics. 2021;22(1):199. Gepubliceerd op 21 maart 2021. doi:10.1186/s12864-021-07537-x(IF:3.969)
[12] Lian C, Yang H, Lan J, et al. Vergelijkende analyse van chloroplastgenomen onthult fylogenetische relaties en intraspecifieke variatie in de medicinale plant Isodon rubescens. PLoS One. 2022;17(4):e0266546. Gepubliceerd op 6 april 2022. doi:10.1371/journal.pone.0266546(IF:3.240)
[13] Diao G, Huang J, Zheng X, et al. Prostaglandine E2 vervult een dubbele rol bij het reguleren van de migratie van dendritische cellen. Int J Mol Med. 2021;47(1):207-218. doi:10.3892/ijmm.2020.4801(IF:3.098)
[14] Bing XL, Zhao DS, Peng CW, Huang HJ, Hong XY. Overeenkomsten en ruimtelijke variaties van bacteriële en schimmelgemeenschappen in populaties van veldrijstplanthoppers (Hemiptera: Delphacidae). Insect Sci. 2020;27(5):947-963. doi:10.1111/1744-7917.12782(IF:2.791)
[15] Li X, Zhou S, Zhang J, Zhou Z, Xiong Q. Directionele veranderingen in de intestinale bacteriële gemeenschap in larven van de zwarte soldatenvlieg (Hermetia illucens). Animals (Basel). 2021;11(12):3475. Gepubliceerd op 6 december 2021. doi:10.3390/ani11123475(IF:2.752)
[16] Yang J, Peng Y, Kong W. Identificatie en volledige genoomsequentie van moerbeicryptisch virus 1. Arch Virol. 2022;167(2):687-690. doi:10.1007/s00705-021-05350-1(IF:2.574)
[17] Chang Y, Xia X, Sui L, et al. Endofytische kolonisatie van entomopathogene schimmels verhoogt de resistentie tegen plantenziekten door de endofytische bacteriële gemeenschap te veranderen. J Basic Microbiol. 2021;61(12):1098-1112. doi:10.1002/taakm.202100494(IF:2.281)
[18] Ding CY, Ma YM, Li B, et al. Identificatie en functionele analyse van differentieel tot expressie gebrachte genen in Myzus persicae (Hemiptera: Aphididae) als reactie op trans-anethol. J Insect Sci. 2022;22(1):3. doi:10.1093/jisesa/ieab094(IF:1.857)
Betaling en beveiliging
Uw betalingsinformatie wordt veilig verwerkt. We slaan geen creditcardgegevens op en hebben geen toegang tot uw creditcardinformatie.
Navraag
Misschien vind je het ook leuk
FAQ
Het product is alleen bedoeld voor onderzoeksdoeleinden en is niet bedoeld voor therapeutisch of diagnostisch gebruik bij mensen of dieren. Producten en inhoud worden beschermd door patenten, handelsmerken en auteursrechten die eigendom zijn van
Voor bepaalde toepassingen zijn mogelijk aanvullende intellectuele eigendomsrechten van derden vereist.