

Beschrijving
GMyc-PCR Mycoplasma Detection Kit gebruikt voornamelijk de PCR-methode om Mycoplasma-infectie van verschillende biologische materialen (zoals celkweek, experimentele dierlijke secreties, dierlijk serum, enz.) te detecteren. Het combineert verschillende voordelen: gevoelig, specifiek, snel en kan direct worden gedetecteerd met celkweeksupernatanten. Dit product detecteert mycoplasma in biologische materialen zoals gekweekte cellen door de PCR-methode. De gebruikte primers zijn ontworpen volgens de geconserveerde regio van de 16S-23S rRNA-sequentie van mycoplasma en versterken alleen specifiek het mycoplasma-DNA, met een hoge detectiegevoeligheid en specificiteit. PCR-amplificatie en elektroforese-analyse duren slechts een paar uur en de bediening is handig en eenvoudig.
Celkweek is een veelvoorkomend experiment in life science-onderzoek. In tegenstelling tot andere veelgebruikte experimentele methoden is celkweek een dynamisch continu proces en cellen reageren vaak op manipulatiefouten of verontreinigingen die vaak abnormale celtoestanden of mediumuiterlijk vertonen. Als het besmet is met mycoplasma, heeft de celmorfologie geen duidelijke verandering en is het gemakkelijk over het hoofd te zien. Het wordt vaak pas ontdekt als de vervuiling zeer ernstig is. Er kunnen honderden mycoplasma's op het besmette celmembraan zitten, deze mycoplasma's concurreren om voedingsstoffen en geven giftige metabolieten af, wat de experimentele resultaten ernstig beïnvloedt.
Uit onderzoek is gebleken dat minstens 20 soorten mycoplasma cellen kunnen besmetten, waarvan de meest voorkomende zijn: orale Mycoplasma (M. orale), Mycoplasma arginine (M. arginini), Mycoplasma hyorhinis (M. hyorhinis), Mycoplasma fermentum (M. fermenten), Mycoplasma hominis (M. hominis), Mycoplasma salivarius (M. speekselklier), Mycoplasma long (M. pulmonis) en Mycoplasma peer (M. pirum). Het mycoplasma-besmettingspercentage van gekweekte cellen varieert van 4% tot 92%. De bronnen van besmetting zijn onder meer de werkomgeving, de operator zelf (sommige mycoplasma's zijn normale flora van het menselijk lichaam), kweekmedium, serum, celkruisbesmetting, experimentele apparatuur en gebruikte besmetting van het oorspronkelijke weefsel of orgaan waaruit cellen zijn bereid.
Het identificeren van de onderliggende oorzaak van problemen tijdens celkweek is een moeilijke en tijdrovende taak, waarbij plotselinge veranderingen verdacht moeten zijn en goede testpraktijken en regelmatige tests op mycoplasma-contaminatie noodzakelijk zijn. Er zijn veel methoden voor de detectie van mycoplasma, zoals directe kweek, DNA-fluorescentiekleuring, ELISA en PCR-methoden.
Functie
- De gebruikte primers zijn ontworpen volgens de geconserveerde regio van de Mycoplasma 16S-23S rRNA-sequentie
- Alleen versterkt mycoplasma-DNA specifiek
- Uitstekende gevoeligheid en specificiteit
- Er konden meer dan 20 mycoplasma-soorten worden gedetecteerd
Sollicitatie
- Mycoplasma-detectie
Componenten
| Componenten nr. | Naam | 40601ES10 (10 testen) | 40601ES20 (20 testen) |
| 40601-A | GMyc-1e PCR-mix | 250 µL | 2×250 µL |
| 40601-B | GMyc-2e PCR-mix | 250 µL | 2×250 µL |
| 40601-C | Positief controlesjabloon N | 20 µL | 20 µL |
[Opmerkingen] 1. Wanneer het gedurende een lange tijd niet wordt gebruikt, kan het worden ingevroren bij -85~-65℃.
2. De PCR-reactie is extreem gevoelig.Om vals-positieve resultaten te voorkomen, wordt er aan het einde van de monstername een positieve controle toegevoegd.
Opslag
Dit product kan 18 maanden bewaard worden bij -25~-15℃. Als het langere tijd niet gebruikt wordt, bewaar het dan uit de buurt van licht.
Cijfers
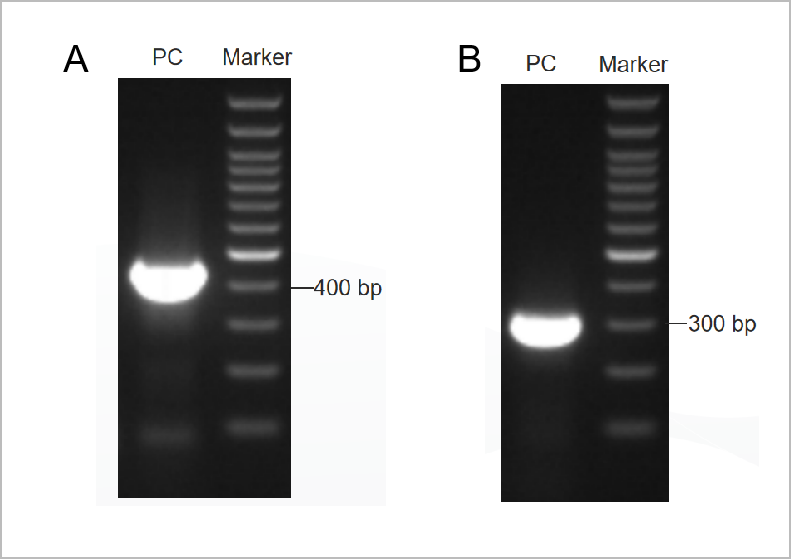
Figuur 1. Het elektroforetogramresultaat van de PCR Mycoplama Test Kit.
Het eerste ronde elektroforetogram werd getoond in Figuur 1A en het tweede ronde elektroforetogram werd getoond in Figuur 1B. (M: 1kb marker, 1-2: positieve controles, 3: de negatieve controle)
[1] Rao XS, Cong XX, Gao XK, et al. AMPK-gemedieerde fosforylering versterkt de auto-inhibitie van TBC1D17 om Rab5-afhankelijke glucoseopname te bevorderen. Cell Death Differ. 2021;28(12):3214-3234. doi:10.1038/s41418-021-00809-9(IF:15.828)
[2] Guo F, Li L, Li J, et al. Single-cell multi-omics sequentiebepaling van vroege muizenembryo's en embryonale stamcellen. Cell Res. 2017;27(8):967-988. doi:10.1038/cr.2017.82(IF:15.606)
[3] Hao Y, He B, Wu L, et al. Nucleaire translocatie van p85β bevordert tumorigenese van PIK3CA-helixdomeinmutante kanker. Nat Commun. 2022;13(1):1974. Gepubliceerd op 13 april 2022. doi:10.1038/s41467-022-29585-x(IF:14.919)
[4] Shu X, Liu M, Lu Z, et al. Genome-wide mapping onthult dat deoxyuridine verrijkt is in het menselijke centromeer-DNA. Nat Chem Biol. 2018;14(7):680-687. doi:10.1038/s41589-018-0065-9(IF:13.843)
[5] Li X, Xiong X, Wang K, et al. Transcriptoom-brede mapping onthult reversibel en dynamisch N(1)-methyladenosine-methyloom. Nat Chem Biol. 2016;12(5):311-316. doi:10.1038/nchembio.2040(IF:12.709)
[6] Sun L, Yang X, Huang X, et al. 2-Hydroxylering van vetzuren onderdrukt colorectale tumorigenese en metastasering via de YAP-transcriptionele as. Cancer Res. 2021;81(2):289-302. doi:10.1158/0008-5472.CAN-20-1517(IF:12.701)
[7] Sun Z, Zhang Z, Wang QQ, Liu JL. Gecombineerde inactivatie van CTPS1 en ATR is synthetisch dodelijk voor MYC-overexpressie kankercellen. Cancer Res. 2022;82(6):1013-1024. doi:10.1158/0008-5472.CAN-21-1707(IF:12.701)
[8] Song J, Zhuang Y, Zhu C, et al. Differentiële rollen van humane PUS10 in miRNA-verwerking en tRNA-pseudouridylatie. Nat Chem Biol. 2020;16(2):160-169. doi:10.1038/s41589-019-0420-5(IF:12.154)
[9] He B, Pan H, Zheng F, et al. Lange niet-coderende RNA LINC00930 bevordert PFKFB3-gemedieerde tumorglycolyse en celproliferatie bij nasofaryngeaal carcinoom. J Exp Clin Cancer Res. 2022;41(1):77. Gepubliceerd 2022 Feb 24. doi:10.1186/s13046-022-02282-9(IF:11.161)
[10] Tang B, Liu BH, Liu ZY, Luo MY, Shi XH, Pang DW. Quantum Dots met een compacte amfifiele zwitterionische coating. ACS Appl Mater Interfaces. 2022;14(24):28097-28104. doi:10.1021/acsami.2c04438(IF:9.229)
[11] Huang C, Zhang Z, Chen L, et al.Acetylering binnen de N- en C-terminale domeinen van Src reguleert verschillende rollen van STAT3-gemedieerde tumorigenese. Cancer Res. 2018;78(11):2825-2838. doi:10.1158/0008-5472.CAN-17-2314(IF:9.130)
[12] Wu X, Yu M, Zhang Z, et al. DDB2 reguleert DNA-replicatie via PCNA-onafhankelijke afbraak van CDT2. Cell Biosci. 2021;11(1):34. Gepubliceerd op 8 feb. 2021. doi:10.1186/s13578-021-00540-5(IF:7.133)
[13] Wang J, Zhang Y, Liu X, Liu H. Optimaliseren van adaptieve therapie op basis van de bereikbaarheid voor tumorresistente subpopulatie. Cancers (Basel). 2021;13(21):5262. Gepubliceerd 2021 okt. 20. doi:10.3390/cancers13215262(IF:6.639)
[14] Feng W, Liu R, Xie X, et al. SUMOylering van α-tubuline is een nieuwe modificatie die de dynamiek van microtubuli reguleert. J Mol Cell Biol. 2021;13(2):91-103. doi:10.1093/jmcb/mjaa076(IF:6.216)
[15] Yu M, Hu X, Yan J, Wang Y, Lu F, Chang J. RIOK2-remmer NSC139021 oefent antitumoreffecten uit op glioblastoom via het induceren van Skp2-gemedieerde celcyclusarrestatie en apoptose. Biomedicines. 2021;9(9):1244. Gepubliceerd op 17 sep 2021. doi:10.3390/biomedicines9091244(IF:6.081)
[16] Ren S, Cai Y, Hu S, et al. Berberine oefent antitumoractiviteit uit in diffuus grootcellig B-cellymfoom door de c-myc/CD47-as te moduleren. Biochem Pharmacol. 2021;188:114576. doi:10.1016/j.bcp.2021.114576(IF:5.858)
[17] Wen F, Sun X, Sun C, et al. TAGLN wordt onderdrukt door TRAF6-gemedieerde proteasomale afbraak in prostaatkankercellen. Mol Cancer Res. 2021;19(7):1113-1122. doi:10.1158/1541-7786.MCR-20-0513(IF:5.852)
[18] Tang B, Sun EZ, Zhang ZL, et al. Sfingomyeline-afgescheiden cholesteroldomein rekruteert formine-bindend eiwit 17 voor het vernauwen van clathrine-gecoate putjes bij de toegang van het influenzavirus. J Virol. 2022;96(5):e0181321. doi:10.1128/JVI.01813-21(IF:5.103)
[19] Hu J, Ren W, Qiu W, et al. Generatie van geïnduceerde pluripotente stamcellijn (XDCMHi001-A) van een patiënt met ankyloserende spondylitis met JAK2-mutatie. Stem Cell Res. 2020;45:101788. doi:10.1016/j.scr.2020.101788(IF:4.495)
[20] Xiao S, Yao X, Ye J, Tian X, Yin Z, Zhou L. Epigenetische modificatie vergemakkelijkt afwijkende expressie van proline synthase PYCR1 bij maagkanker [online gepubliceerd vóór druk, 30 mei 2022]. Biochim Biophys Acta Gene Regul Mech. 2022;1865(6):194829. doi:10.1016/j.bbagrm.2022.194829(IF:4.490)
[21] Wang J, Zhang Y, Liu X, Liu H. Is de vaste periodieke behandeling effectief voor het tumorsysteem zonder volledige informatie?. Cancer Manag Res. 2021;13:8915-8928. Gepubliceerd 30 nov. 2021. doi:10.2147/CMAR.S339787(IF:3.989)
[22] Yang X, Ren S, Rehman ZU, et al. Moleculaire karakterisering, expressie en functionele identificatie van TANK-bindende kinase 1 (TBK1) van de koe (Bos taurus) en de geit (Capra hircus). Dev Comp Immunol. 2022;133:104444. doi:10.1016/j.dci.2022.104444(IF:3.636)
[23] Zheng D, Chang X, Liu Y, et al. 2-Methoxy-5((3,4,5-trimethosyfenyl)seleninyl)fenol keert EGF-geïnduceerde celmigratie en invasie om door downregulatie van MDM2 in borstkankercellijnen. Cancer Biol Ther. 2019;20(4):513-523. doi:10.1080/15384047.2018.1537578(IF:3.373)
[24] Xu F, Zhang S, Liu Z, et al. TEX9 en eIF3b werken functioneel synergetisch samen om de progressie van plaveiselcelcarcinoom van de slokdarm te bevorderen. BMC Cancer. 2019;19(1):875. Gepubliceerd op 3 sep 2019. doi:10.1186/s12885-019-6071-9(IF:2.933)
[25] Pan H, Sun L, Wang W, et al. Serum lange niet-coderende RNA LOC553103 als niet-specifieke diagnostische en prognostische biomarker voor veel voorkomende soorten menselijke kanker. Clin Chim Acta. 2020;508:69-76. doi:10.1016/j.cca.2020.05.017(IF:2.615)
[26] Li N, Lin SM, Li Y, Zon J, Zhang L, Chen M.Een geïnduceerde pluripotente stamcellijn (GZHMCi004-A) afgeleid van een foetus met heterozygote G380R-mutatie in het FGFR3-gen dat achondroplasie veroorzaakt. Stem Cell Res. 2021;53:102322. doi:10.1016/j.scr.2021.102322(IF:2.020)
[27] Luo Q, Wei C, Long Y, et al. Generatie van een ELTD1 knockout humane embryonale stamcellijn door het iCRISPR/Cas9-systeem. Stem Cell Res. 2021;53:102350. doi:10.1016/j.scr.2021.102350(IF:2.020)
[28] Liu YQ, Ling TW, Wang HY, Yang YH, Song WJ, Wang TC. Generatie van een integratievrije geïnduceerde pluripotente stamcellijn (LZUSHI001-A) van een epileptische patiënt met DGKG-mutatie. Stem Cell Res. 2022;61:102768. doi:10.1016/j.scr.2022.102768(IF:2.020)
[29] Chen M, Lin SM, Li N, Li Y, Li Y, Zhang L. Een geïnduceerde pluripotente stamcellijn (GZHMCi003-A) afgeleid van een foetus met exon 3 heterozygote deletie in het RUNX2-gen dat cleidocraniale dysplasie veroorzaakt. Stem Cell Res. 2021;51:102166. doi:10.1016/j.scr.2021.102166(IF:2.020)
[30] Xu Y, Wang X, Qiu T, et al. Generatie van een geïnduceerde pluripotente stamcellijn (FDCHI007-A) afgeleid van een patiënt met ontwikkelings- en epileptische encefalopathie type 31 die heterozygote c.545C draagt > Een mutatie in het DNM1-gen. Stem Cell Res. 2022;60:102709. doi:10.1016/j.scr.2022.102709(IF:2.020)
[31] Fan T, He J, Wang Y, Yu J, Sun W. Generatie van een geïnduceerde pluripotente stamcellijn (FDCHi006-A) van een 7-jarig meisje met centrale vroegtijdige puberteit. Stem Cell Res. 2021;56:102542. doi:10.1016/j.scr.2021.102542(IF:2.020)
[32] Gong X, Zheng Z, Yang T, Zheng H, Xiao X, Jia N. Generatie van een isogene gen-gecorrigeerde iPSC-lijn (OGHFUi001-A-1) van een patiënt met type 1 vroege infantiele epileptische encefalopathie (EIEE1) met een hemizygote R330L-mutatie in het ARX-gen. Stem Cell Res. 2022;60:102693. doi:10.1016/j.scr.2022.102693(IF:2.020)
[33] Jia N, Gong X, Chen J, et al. Generatie van een geïnduceerde pluripotente stamcellijn (OGHFUi001-A) uit een type 1 vroege infantiele epileptische encefalopathie met ARX-mutatie. Stem Cell Res. 2021;53:102367. doi:10.1016/j.scr.2021.102367(IF:2.020)
[34] Zhu W, Zhou Y, Wang Q, et al. Generatie van een door de mens geïnduceerde pluripotente stamcellijn (iPSC) uit huidfibroblasten van een patiënt die een E363Q-mutatie in het PSEN1-gen draagt. Stem Cell Res. 2022;61:102769. doi:10.1016/j.scr.2022.102769(IF:2.020)
[35] Luo F, Long K, Li X, et al. Een tekort aan LRRC8A verzwakt door hypoxie geïnduceerde necrose in 3T3-L1-cellen. Biosci Biotechnol Biochem. 2020;84(6):1139-1145. doi:10.1080/09168451.2020.1730689(IF:1.516)
Betaling en beveiliging
Uw betalingsinformatie wordt veilig verwerkt. We slaan geen creditcardgegevens op en hebben geen toegang tot uw creditcardinformatie.
Navraag
Misschien vind je het ook leuk
FAQ
Het product is alleen bedoeld voor onderzoeksdoeleinden en is niet bedoeld voor therapeutisch of diagnostisch gebruik bij mensen of dieren. Producten en inhoud worden beschermd door patenten, handelsmerken en auteursrechten die eigendom zijn van
Voor bepaalde toepassingen zijn mogelijk aanvullende intellectuele eigendomsrechten van derden vereist.