





Описание
Бисеры для выбора ДНК Hieff NGS™ готовятся на основе принципа SPRI (твердофазная обратная иммобилизация) и применяются для очистки ДНК и выбора размера во время подготовки библиотек секвенирования следующего поколения (NGS). Бисеры для выбора ДНК Hieff NGS™ совместимы с различными наборами для подготовки библиотек ДНК и РНК и являются хорошей альтернативой AMЧистые бусины.
Компоненты
| Номер компонента | Имя | 12601ES08 | 12601ES56 | 12601ES75 |
| 12601 | Бисеры для отбора ДНК Hieff NGS™ | 5 мл | 60 мл | 450 мл |
Технические характеристики
| Линия продуктов | Бусины для очистки и отбора ДНК |
| Исходный материал | ДНК |
| Совместимость | ДНК |
| Технология изоляции | Магнитная бусина |
| Тип конечного продукта | ДНК |
| Для использования с (приложение) | ДНК целан вверх, выбор размера ДНК |
Доставка и хранение
Бусины поставляются с пакетами со льдом и могут храниться при температуре от 2°C до 8°C в течение одного года.
Инструкции
- 1. Подготовка
Перед использованием выдержите селективные шарики при комнатной температуре не менее 30 минут.
- 2. Выбор размера ДНК
Последовательность операций выбора размера показана на рисунке 1, а протокол выглядит следующим образом.
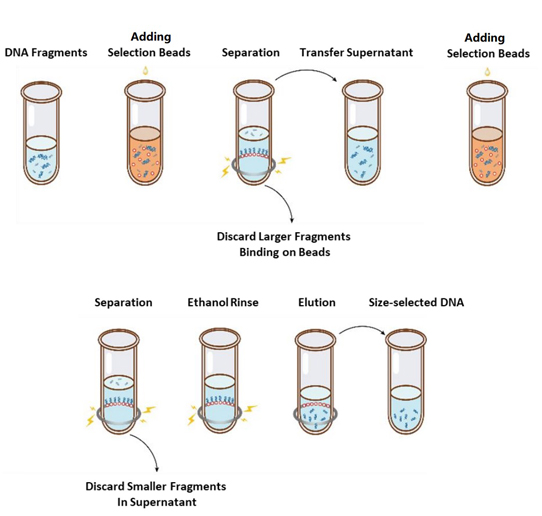
Рисунок 1. Блок-схема выбора размера ДНК
2.1 Перед каждым использованием тщательно перемешивайте гранулы путем встряхивания или пипетирования.
2.2 Добавьте первый раунд отборочных шариков к образцу (см. Таблицу 1). Тщательно перемешайте путем вортексирования или пипетирования вверх и вниз не менее 10 раз.
2.3 Инкубируйте при комнатной температуре в течение 5 мин.
2.4 Быстро центрифугируйте пробирку и поместите ее на магнитный штатив. Когда раствор станет прозрачным (примерно через 5 мин), перенесите супернатант в новую пробирку для ПЦР.
2.5 Добавьте вторую порцию отборочных шариков к образцу из шага 2.4 в соответствии с таблицей 1. Тщательно перемешайте путем встряхивания или пипетирования вверх и вниз не менее 10 раз.
2.6 Инкубируйте при комнатной температуре в течение 5 мин.
2.7 Быстро центрифугируйте пробирку и поместите ее на магнитный штатив. Когда раствор станет прозрачным (примерно через 5 мин), аспирируйте надосадочную жидкость и выбросьте ее.
2.8 Поместите пробирку в магнитный штатив и добавьте 200 мкл свежеприготовленного 80% этанола, не трогая гранулы, инкубируйте при комнатной температуре в течение 30 сек. Аспирируйте этанол и выбросьте.
2.9 Повторите шаг 2.8 один раз, чтобы выполнить в общей сложности две стирки.
2.10 Удалите остаточный этанол с помощью наконечников пипеток объемом 10 мкл. Держите пробирку в магнитном штативе, высушите на воздухе селективные бусины с открытой крышкой до тех пор, пока не появятся трещины (около 5 мин).
Примечание: Не пересушивайте гранулы для отбора проб. Это может привести к снижению эффективности восстановления ДНК-мишени.
2.11 Снимите пробирку с магнитного штатива. Добавьте соответствующее количество ddH2O (≥20 мкл) и тщательно перемешайте путем вортексирования или пипетирования вверх и вниз не менее 10 раз.
2.12 Инкубируйте при комнатной температуре в течение 5 мин.
Быстро опустите пробирку и поместите ее на магнитный штатив. Когда раствор станет прозрачным (примерно через 5 минут), перенесите 20 мкл супернатанта в новую пробирку.
- 3.Рекомендуемые условия для выбора размера ДНК
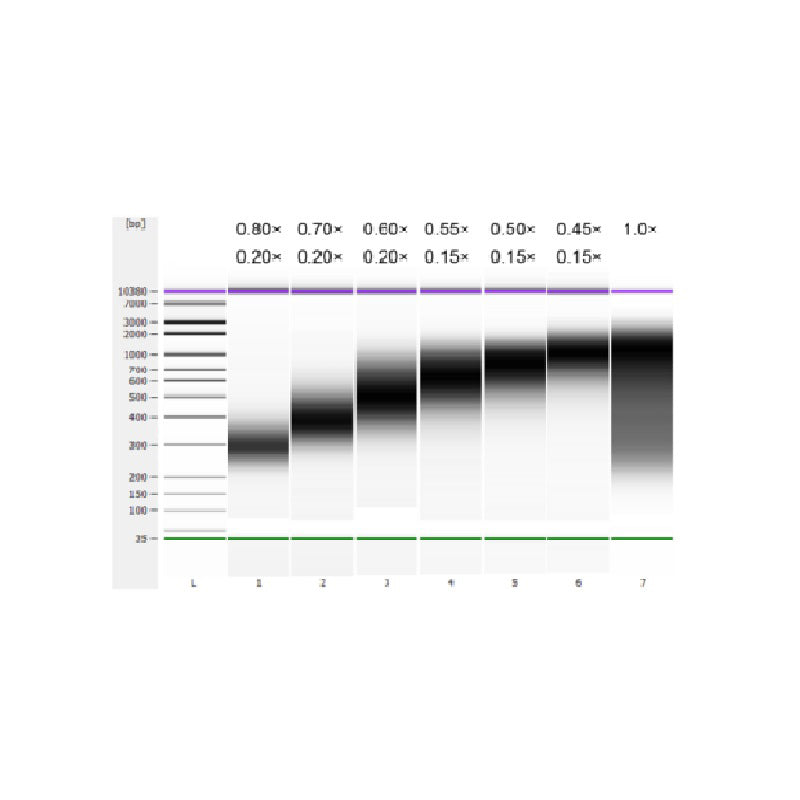
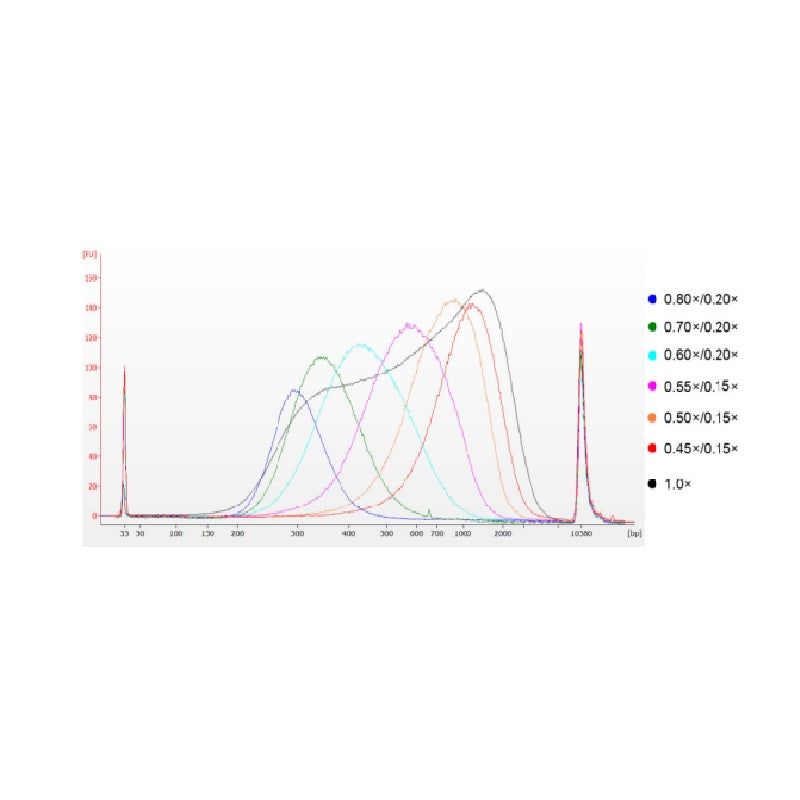
ДНК тимуса теленка была фрагментирована с помощью ультразвука для получения фрагмента размером 100–1000 п.н., и были проведены два раунда отбора размера в соответствии с таблицей 1. Результаты были проанализированы с помощью биоанализатора Agilent 2100 (рисунок 2).
Таблица 1. Рекомендуемые условия для выбора размера ДНК
| Длина фрагмента ДНК | 250-350 п.н. | 320-420 п.н. | 450-550 п.н. | 550-700 п.н. | 700-900 п.н. | 800-1000 п.н. |
| Соотношение бусин: ДНК для 1-го раунда | 0,80× | 0,70× | 0,60× | 0,55× | 0,50× | 0,45× |
| Соотношение бусин: ДНК для 2-го Круглый | 0,20× | 0,20× | 0,20× | 0,15× | 0,15× | 0,15× |
Примечание: «×» в таблице указывает объем образца ДНК. Например, если длина вставки библиотеки составляет 250 п.н., а объем образца ДНК составляет 100 мкл, объем магнитных частиц, используемых в первом раунде сортировки, составляет 0,80×100 мкл=80 мкл; объем магнитных частиц, используемых во втором раунде сортировки, составляет 0,20×100 мкл=20 мкл.
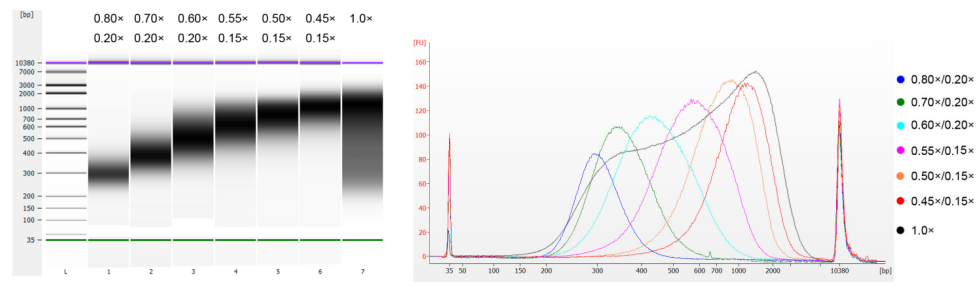
Рисунок 2. Электрофореграмма высокочувствительного ДНК-чипа Agilent 2100
Примечания:
1. Для вашей безопасности и здоровья, пожалуйста, надевайте лабораторный халат и одноразовые перчатки во время работы.

Цитируется из «Специфическая интеграция последовательности каспозазой семейства 1 из Candidatus Nitrosopumilus koreensis» AR1. Nucleic Acids Res. 2021;49(17):9938-9952. doi:10.1093/nar/gkab725"

Цитируется из «Недавнее заражение Wolbachia изменяет микробные сообщества в популяциях диких Laodelphax striatellus. Microbiome. 2020;8(1):104. Опубликовано 2 июля 2020 г. doi:10.1186/s40168-020-00878-x»
[1] Ван X, Юань Q, Чжан В и др. Последовательность-специфическая интеграция каспозазой семейства 1 из Candidatus Nitrosopumilus koreensis AR1. Nucleic Acids Res. 2021;49(17):9938-9952. doi:10.1093/nar/gkab725(IF:16.971)
[2] Duan XZ, Sun JT, Wang LT и др. Недавнее заражение Wolbachia изменяет микробные сообщества в популяциях диких Laodelphax striatellus. Microbiome. 2020;8(1):104. Опубликовано 2 июля 2020 г. doi:10.1186/s40168-020-00878-x(IF:11.607)
[3] Сонг Б., Альматрафи Е., Санг Ф. и др.Управление осадком, обработанным по методу Фентона, с помощью биоугля и компоста из овечьего навоза: влияние на эволюционные характеристики бактериального сообщества. J Environ Manage. 2022;316:115218. doi:10.1016/j.jenvman.2022.115218(IF:6.789)
[4] Хуан С., Мэй К., Лу Л. и др. Язвенный колит в ответ на трансплантацию фекальной микробиоты посредством модуляции кишечной микробиоты и баланса клеток Th17/Treg. Клетки. 2022;11(11):1851. Опубликовано 5 июня 2022 г. doi:10.3390/cells11111851(IF:6.600)
[5] Ghosh S, Yang X, Wang L, Zhang C, Zhao L. Активная фаза пребиотического питания изменяет микробиоту кишечника, вызывает независимое от веса облегчение гепатостеатоза и уровня сывороточного холестерина у мышей, получавших диету с высоким содержанием жиров. Comput Struct Biotechnol J. 2020;19:448-458. Опубликовано 24 декабря 2020 г. doi:10.1016/j.csbj.2020.12.011(IF:6.018)
[6] Гао X, Ю Б, Ю Дж и др. Профилирование развития переваривания пищевых углеводов у поросят. Front Microbiol. 2022;13:896660. Опубликовано 29 апреля 2022 г. doi:10.3389/fmicb.2022.896660(IF:5.640)
[7] Ли П., Чжан И., Янь Ф., Чжоу С. Характеристики бактериофага vB_Kox_ZX8, выделенного из клинической Klebsiella oxytoca, и его терапевтический эффект на бактериемию у мышей. Front Microbiol. 2021;12:763136. Опубликовано 3 декабря 2021 г. doi:10.3389/fmicb.2021.763136(IF:5.640)
[8] Линь З., Луо П., Хуан Д., У И., Ли Ф., Лю Х. Стратегия анализа токсичности акриламида в модели Saccharomyces cerevisiae на основе мультиомики. Chem Biol Interact. 2021;349:109682. doi:10.1016/j.cbi.2021.109682(IF:5.194)
[9] Сунь X, Lv W, Ван Y и др. Ген Mrgprb2 играет роль в анафилактоидных реакциях, вызванных инъекцией хауттюйнии сердцевидной. Дж Этнофармакол. 2022;289:115053. doi:10.1016/j.jep.2022.115053(ЕСЛИ:4.360)
[10] Ma H, Lai B, Zan C, Di X, Zhu X, Wang K. GLO1 способствует лекарственной устойчивости Escherichia coli посредством индукции PER-типа β-лактамаз расширенного спектра. Infect Drug Resist. 2022;15:1573-1586. Опубликовано 5 апреля 2022 г. doi:10.2147/IDR.S358578(IF:4.003)
[11] Zhong Y, Zhao W, Tang Z и др. Сравнительный транскриптомный анализ различных стадий развития яичника красного болотного рака Procambarus clarkii. BMC Genomics. 2021;22(1):199. Опубликовано 21 марта 2021 г. doi:10.1186/s12864-021-07537-x(IF:3.969)
[12] Lian C, Yang H, Lan J и др. Сравнительный анализ геномов хлоропластов выявляет филогенетические связи и внутривидовые различия в лекарственном растении Isodon rubescens. PLoS One. 2022;17(4):e0266546. Опубликовано 6 апреля 2022 г. doi:10.1371/journal.pone.0266546(IF:3.240)
[13] Дяо Г., Хуан Дж., Чжэн Х. и др. Простагландин E2 выполняет двойную роль в регуляции миграции дендритных клеток. Int J Mol Med. 2021;47(1):207-218. doi:10.3892/ijmm.2020.4801(IF:3.098)
[14] Bing XL, Zhao DS, Peng CW, Huang HJ, Hong XY. Сходства и пространственные вариации бактериальных и грибковых сообществ в популяциях полевых рисовых цикад (Hemiptera: Delphacidae). Insect Sci. 2020;27(5):947-963. doi:10.1111/1744-7917.12782(IF:2.791)
[15] Ли X, Чжоу С, Чжан Дж, Чжоу З, Сюн Ц. Направленные изменения в кишечном бактериальном сообществе у личинок черной львинки (Hermetia illucens). Животные (Базель). 2021;11(12):3475. Опубликовано 6 декабря 2021 г. doi:10.3390/ani11123475(IF:2.752)
[16] Yang J, Peng Y, Kong W. Идентификация и полная последовательность генома криптического вируса шелковицы 1. Arch Virol. 2022;167(2):687-690. doi:10.1007/s00705-021-05350-1(IF:2.574)
[17] Чанг И., Ся Х., Суй Л. и др. Эндофитная колонизация энтомопатогенных грибов повышает устойчивость растений к болезням за счет изменения эндофитного бактериального сообщества. J Basic Microbiol. 2021;61(12):1098-1112. doi:10.1002/jobm.202100494(IF:2.281)
[18] Ding CY, Ma YM, Li B и др. Идентификация и функциональный анализ дифференциально экспрессируемых генов у Myzus persicae (Hemiptera: Aphididae) в ответ на транс-анетол. J Insect Sci. 2022;22(1):3. doi:10.1093/jisesa/ieab094(IF:1.857)
Оплата и безопасность
Ваша платежная информация обрабатывается надежно. Мы не храним данные кредитной карты и не имеем доступа к информации вашей кредитной карты.
Расследование
Вам также может понравиться
Часто задаваемые вопросы
Продукт предназначен только для исследовательских целей и не предназначен для терапевтического или диагностического использования на людях или животных. Продукты и содержимое защищены патентами, товарными знаками и авторскими правами, принадлежащими Yeasen Biotechnology. Символы товарных знаков указывают на страну происхождения, а не обязательно на регистрацию во всех регионах.
Для некоторых приложений могут потребоваться дополнительные права интеллектуальной собственности третьих лиц.
Йесен привержен этической науке, полагая, что наши исследования должны затрагивать важнейшие вопросы, обеспечивая при этом безопасность и соблюдение этических стандартов.