

Описание
Набор для обнаружения микоплазмы GMyc-PCR в основном использует метод ПЦР для обнаружения микоплазменной инфекции в различных биологических материалах (таких как клеточная культура, выделения экспериментальных животных, сыворотка животных и т. д.). Он сочетает в себе несколько преимуществ: чувствительность, специфичность, скорость и может быть обнаружен непосредственно с супернатантами клеточной культуры. Этот продукт обнаруживает микоплазму в биологических материалах, таких как культивируемые клетки, методом ПЦР. Используемые праймеры разработаны в соответствии с консервативной областью последовательности рРНК 16S-23S микоплазмы и специфически амплифицируют только ДНК микоплазмы с высокой чувствительностью и специфичностью обнаружения. Амплификация ПЦР и анализ электрофореза занимают всего несколько часов, а операция удобна и проста.
Культивирование клеток является обычным экспериментом в исследованиях в области естественных наук. В отличие от других широко используемых экспериментальных методов, культивирование клеток является динамическим непрерывным процессом, и клетки часто реагируют на ошибки манипуляции или загрязняющие вещества, которые часто демонстрируют ненормальные состояния клеток или внешний вид среды. Если она загрязнена микоплазмой, морфология клеток не имеет очевидных изменений, и ее легко пропустить. Ее часто не обнаруживают, пока загрязнение не станет очень серьезным. На загрязненной клеточной мембране могут быть сотни микоплазм, эти микоплазмы конкурируют за питательные вещества и выделяют токсичные метаболиты, серьезно влияя на результаты эксперимента.
Исследования показали, что по крайней мере 20 видов микоплазм могут заражать клетки, среди которых наиболее распространенными являются: оральная микоплазма (M. orale), микоплазма аргинина (М. аргинини), Mycoplasma hyorhinis (М. hyorhinis), Mycoplasma fermentum (М. fermentans), Mycoplasma hominis (М. хоминис), Mycoplasma salivarius (М. слюнной железы), микоплазма легких (М. легочная) и микоплазма груши (М. пирум). Уровень заражения культивируемых клеток микоплазмами составляет от 4% до 92%. Источниками заражения являются рабочая среда, сам оператор (некоторые микоплазмы являются нормальной флорой человеческого организма), питательная среда, сыворотка, перекрестное загрязнение клеток, экспериментальное оборудование и использованное загрязнение исходной ткани или органа, из которых были получены клетки.
Выявление основной причины проблем во время культивирования клеток — сложная и трудоемкая задача, где следует подозревать любые внезапные изменения, и необходимы надлежащие методы тестирования и регулярные проверки на загрязнение микоплазмой. Существует много методов обнаружения микоплазмы, таких как прямая культура, флуоресцентное окрашивание ДНК, ИФА и методы ПЦР.
Особенность
- Использованные праймеры были разработаны в соответствии с консервативной областью последовательности рРНК Mycoplasma 16S-23S.
- Только амплифицированная ДНК микоплазмы специфически
- Отличная чувствительность и специфичность
- Удалось обнаружить более 20 видов микоплазм
Приложение
- Обнаружение микоплазмы
Компоненты
| Номер компонента | Имя | 40601ES10 (10 анализов) | 40601ES20 (20 анализов) |
| 40601-А | GMyc-1st ПЦР-смесь | 250 мкл | 2×250 мкл |
| 40601-Б | GMyc-2nd ПЦР-смесь | 250 мкл | 2×250 мкл |
| 40601-С | Положительный контрольный шаблон N | 20 мкл | 20 мкл |
[Примечания] 1. Если устройство не используется в течение длительного времени, его можно хранить в замороженном виде при температуре -85~-65 ℃.
2. Реакция ПЦР чрезвычайно чувствительна.Для предотвращения ложноположительных результатов в конце добавления образцов добавляется положительный контроль.
Хранилище
Этот продукт может храниться при температуре -25~-15℃ в течение 18 месяцев. Если он не используется в течение длительного времени, пожалуйста, держите его вдали от света.
Цифры
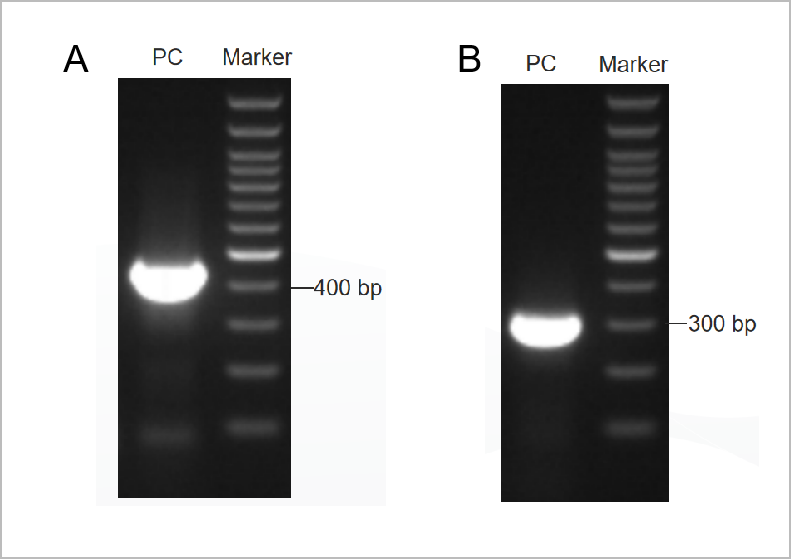
Рисунок 1. Результат электрофореграммы набора для ПЦР-теста на микоплазму.
Электрофореграмма первого раунда показана на рисунке 1А, а электрофореграмма второго раунда показана на рисунке 1Б. (М: маркер 1 кб, 1-2: положительный контроль, 3: отрицательный контроль)
[1] Rao XS, Cong XX, Gao XK и др. Фосфорилирование, опосредованное AMPK, усиливает аутоингибирование TBC1D17, способствуя Rab5-зависимому поглощению глюкозы. Cell Death Differ. 2021;28(12):3214-3234. doi:10.1038/s41418-021-00809-9(IF:15.828)
[2] Го Ф., Ли Л., Ли Дж. и др. Одноклеточное мультиомное секвенирование ранних эмбрионов мыши и эмбриональных стволовых клеток. Cell Res. 2017;27(8):967-988. doi:10.1038/cr.2017.82(IF:15.606)
[3] Хао И., Хе Б., У Л. и др. Ядерная транслокация p85β способствует возникновению опухолей при раке, вызванном мутацией спирального домена PIK3CA. Nat Commun. 2022;13(1):1974. Опубликовано 13 апреля 2022 г. doi:10.1038/s41467-022-29585-x(IF:14.919)
[4] Шу X, Лю М, Лу З и др. Картирование генома показывает, что дезоксиуридин обогащен в человеческой центромерной ДНК. Nat Chem Biol. 2018;14(7):680-687. doi:10.1038/s41589-018-0065-9(IF:13.843)
[5] Ли X, Сюн X, Ван К и др. Картирование транскриптома выявляет обратимый и динамический метилом N(1)-метиладенозина. Nat Chem Biol. 2016;12(5):311-316. doi:10.1038/nchembio.2040(IF:12.709)
[6] Sun L, Yang X, Huang X и др. 2-Гидроксилирование жирных кислот подавляет колоректальный опухолегенез и метастазирование через транскрипционную ось YAP. Cancer Res. 2021;81(2):289-302. doi:10.1158/0008-5472.CAN-20-1517(IF:12.701)
[7] Sun Z, Zhang Z, Wang QQ, Liu JL. Комбинированная инактивация CTPS1 и ATR является синтетически летальной для раковых клеток, сверхэкспрессирующих MYC. Cancer Res. 2022;82(6):1013-1024. doi:10.1158/0008-5472.CAN-21-1707(IF:12.701)
[8] Song J, Zhuang Y, Zhu C и др. Дифференциальные роли человеческого PUS10 в процессинге miRNA и псевдоуридилировании tRNA. Nat Chem Biol. 2020;16(2):160-169. doi:10.1038/s41589-019-0420-5(IF:12.154)
[9] He B, Pan H, Zheng F и др. Длинная некодирующая РНК LINC00930 способствует PFKFB3-опосредованному опухолевому гликолизу и пролиферации клеток при носоглоточной карциноме. J Exp Clin Cancer Res. 2022;41(1):77. Опубликовано 24 февраля 2022 г. doi:10.1186/s13046-022-02282-9(IF:11.161)
[10] Тан Б., Лю Б. Х., Лю З. И., Луо М. Ю., Ши Х. Х., Пан Д. В. Квантовые точки с компактным амфифильным цвиттерионным покрытием. ACS Appl Mater Interfaces. 2022;14(24):28097-28104. doi:10.1021/acsami.2c04438(IF:9.229)
[11] Хуан С., Чжан Цз., Чен Л. и др.Ацетилирование в пределах N- и C-концевых доменов Src регулирует различные роли STAT3-опосредованного опухолеобразования. Cancer Res. 2018;78(11):2825-2838. doi:10.1158/0008-5472.CAN-17-2314(IF:9.130)
[12] Wu X, Yu M, Zhang Z и др. DDB2 регулирует репликацию ДНК посредством PCNA-независимой деградации CDT2. Cell Biosci. 2021;11(1):34. Опубликовано 8 февраля 2021 г. doi:10.1186/s13578-021-00540-5(IF:7.133)
[13] Ван Дж., Чжан И., Лю Х., Лю Х. Оптимизация адаптивной терапии на основе доступности субпопуляции, устойчивой к опухолям. Рак (Базель). 2021;13(21):5262. Опубликовано 20 октября 2021 г. doi:10.3390/cancers13215262(IF:6.639)
[14] Feng W, Liu R, Xie X и др. SUMO-илирование α-тубулина — новая модификация, регулирующая динамику микротрубочек. J Mol Cell Biol. 2021;13(2):91-103. doi:10.1093/jmcb/mjaa076(IF:6.216)
[15] Yu M, Hu X, Yan J, Wang Y, Lu F, Chang J. Ингибитор RIOK2 NSC139021 оказывает противоопухолевое действие на глиобластому посредством индукции остановки клеточного цикла, опосредованной Skp2, и апоптоза. Biomedicines. 2021;9(9):1244. Опубликовано 17 сентября 2021 г. doi:10.3390/biomedicines9091244(IF:6.081)
[16] Ren S, Cai Y, Hu S и др. Берберин оказывает противоопухолевое действие при диффузной В-клеточной лимфоме, модулируя ось c-myc/CD47. Biochem Pharmacol. 2021;188:114576. doi:10.1016/j.bcp.2021.114576(IF:5.858)
[17] Вэнь Ф., Сан Х., Сан К. и др. TAGLN подавляется протеасомной деградацией, опосредованной TRAF6, в клетках рака простаты. Mol Cancer Res. 2021;19(7):1113-1122. doi:10.1158/1541-7786.MCR-20-0513(IF:5.852)
[18] Тан Б., Сан Э.З., Чжан З.Л. и др. Сфингомиелин-секвестрированный домен холестерина привлекает формин-связывающий белок 17 для сужения покрытых клатрином ямок при проникновении вируса гриппа. J Virol. 2022;96(5):e0181321. doi:10.1128/JVI.01813-21(IF:5.103)
[19] Ху Дж., Рен В., Цю В. и др. Генерация линии индуцированных плюрипотентных стволовых клеток (XDCMHi001-A) от пациента с анкилозирующим спондилитом с мутацией JAK2. Stem Cell Res. 2020;45:101788. doi:10.1016/j.scr.2020.101788(IF:4.495)
[20] Xiao S, Yao X, Ye J, Tian X, Yin Z, Zhou L. Эпигенетическая модификация способствует аберрантной экспрессии пролинсинтазы PYCR1 при раке желудка [опубликовано онлайн до выхода в печать, 30 мая 2022 г.]. Biochim Biophys Acta Gene Regul Mech. 2022;1865(6):194829. doi:10.1016/j.bbagrm.2022.194829(IF:4.490)
[21] Ван Дж., Чжан И., Лю Х., Лю Х. Эффективно ли фиксированное периодическое лечение для опухолевой системы без полной информации? Cancer Manag Res. 2021;13:8915-8928. Опубликовано 30 ноября 2021 г. doi:10.2147/CMAR.S339787(IF:3.989)
[22] Yang X, Ren S, Rehman ZU и др. Молекулярная характеристика, экспрессия и функциональная идентификация TANK-связывающей киназы 1 (TBK1) коровы (Bos taurus) и козы (Capra hircus). Dev Comp Immunol. 2022;133:104444. doi:10.1016/j.dci.2022.104444(IF:3.636)
[23] Чжэн Д., Чанг Х., Лю И. и др. 2-Метокси-5((3,4,5-триметосифенил)селенинил) фенол обращает вспять миграцию и инвазию клеток, вызванную EGF, посредством снижения регуляции MDM2 в клеточных линиях рака молочной железы. Cancer Biol Ther. 2019;20(4):513-523. doi:10.1080/15384047.2018.1537578(IF:3.373)
[24] Xu F, Zhang S, Liu Z и др. TEX9 и eIF3b функционально взаимодействуют, способствуя прогрессированию плоскоклеточного рака пищевода. BMC Cancer. 2019;19(1):875. Опубликовано 3 сентября 2019 г. doi:10.1186/s12885-019-6071-9(IF:2.933)
[25] Pan H, Sun L, Wang W и др. Длинная некодирующая РНК сыворотки LOC553103 как неспецифический диагностический и прогностический биомаркер для распространенных типов рака человека. Clin Chim Acta. 2020;508:69-76. doi:10.1016/j.cca.2020.05.017(IF:2.615)
[26] Ли Н, Линь С.М., Ли Ю, Сунь Дж, Чжан Л, Чен М.Индуцированная плюрипотентная линия стволовых клеток (GZHMCi004-A), полученная от плода с гетерозиготной мутацией G380R в гене FGFR3, вызывающей ахондроплазию. Stem Cell Res. 2021;53:102322. doi:10.1016/j.scr.2021.102322(IF:2.020)
[27] Luo Q, Wei C, Long Y и др. Создание линии эмбриональных стволовых клеток человека с нокаутом ELTD1 с помощью системы iCRISPR/Cas9. Stem Cell Res. 2021;53:102350. doi:10.1016/j.scr.2021.102350(IF:2.020)
[28] Liu YQ, Ling TW, Wang HY, Yang YH, Song WJ, Wang TC. Генерация линии индуцированных плюрипотентных стволовых клеток без интеграции (LZUSHI001-A) от пациента с эпилепсией с мутацией DGKG. Stem Cell Res. 2022;61:102768. doi:10.1016/j.scr.2022.102768(IF:2.020)
[29] Chen M, Lin SM, Li N, Li Y, Li Y, Zhang L. Индуцированная плюрипотентная линия стволовых клеток (GZHMCi003-A), полученная от плода с гетерозиготной делецией экзона 3 в гене RUNX2, вызывающей ключично-черепную дисплазию. Stem Cell Res. 2021;51:102166. doi:10.1016/j.scr.2021.102166(IF:2.020)
[30] Xu Y, Wang X, Qiu T и др. Создание линии индуцированных плюрипотентных стволовых клеток (FDCHI007-A), полученных от пациента с энцефалопатией развития и эпилептической энцефалопатией типа 31, несущей гетерозиготный c.545C > Мутация в гене DNM1. Stem Cell Res. 2022;60:102709. doi:10.1016/j.scr.2022.102709(IF:2.020)
[31] Fan T, He J, Wang Y, Yu J, Sun W. Генерация линии индуцированных плюрипотентных стволовых клеток (FDCHi006-A) от 7-летней девочки с центральным преждевременным половым созреванием. Stem Cell Res. 2021;56:102542. doi:10.1016/j.scr.2021.102542(IF:2.020)
[32] Gong X, Zheng Z, Yang T, Zheng H, Xiao X, Jia N. Генерация изогенной генно-скорректированной линии iPSC (OGHFUi001-A-1) от пациента с ранней детской эпилептической энцефалопатией 1 типа (EIEE1) с гемизиготной мутацией R330L в гене ARX. Stem Cell Res. 2022;60:102693. doi:10.1016/j.scr.2022.102693(IF:2.020)
[33] Jia N, Gong X, Chen J и др. Генерация линии индуцированных плюрипотентных стволовых клеток (OGHFUi001-A) из ранней детской эпилептической энцефалопатии 1 типа с мутацией ARX. Stem Cell Res. 2021;53:102367. doi:10.1016/j.scr.2021.102367(IF:2.020)
[34] Чжу В., Чжоу И., Ван К. и др. Создание линии человеческих индуцированных плюрипотентных стволовых клеток (iPSC) из фибробластов кожи пациента, несущего мутацию E363Q в гене PSEN1. Stem Cell Res. 2022;61:102769. doi:10.1016/j.scr.2022.102769(IF:2.020)
[35] Луо Ф., Лонг К., Ли Х. и др. Дефицит LRRC8A ослабляет некроз, вызванный гипоксией, в клетках 3T3-L1. Biosci Biotechnol Biochem. 2020;84(6):1139-1145. doi:10.1080/09168451.2020.1730689(IF:1.516)
Оплата и безопасность
Ваша платежная информация обрабатывается надежно. Мы не храним данные кредитной карты и не имеем доступа к информации вашей кредитной карты.
Расследование
Вам также может понравиться
Часто задаваемые вопросы
Продукт предназначен только для исследовательских целей и не предназначен для терапевтического или диагностического использования на людях или животных. Продукты и содержимое защищены патентами, товарными знаками и авторскими правами, принадлежащими Yeasen Biotechnology. Символы товарных знаков указывают на страну происхождения, а не обязательно на регистрацию во всех регионах.
Для некоторых приложений могут потребоваться дополнительные права интеллектуальной собственности третьих лиц.
Йесен привержен этической науке, полагая, что наши исследования должны затрагивать важнейшие вопросы, обеспечивая при этом безопасность и соблюдение этических стандартов.