


Beskrivning
Hieff Trans™ Liposomal Transfection Reagent är ett mångsidigt liposomtransfektionsreagens, lämpligt för DNA-, RNA- och oligonukleotidtransfektion, med hög transfektionseffektivitet för de flesta eukaryota celler. Dess unika formel gör att det kan tillsättas direkt till mediet, och närvaron av serum påverkar inte transfektionseffektiviteten, vilket minskar skadorna på celler som orsakas av serumavlägsnande. Det finns inget behov av att ta bort nukleinsyra-Hieff Trans™-komplexet eller ersätta det med ett nytt medium efter transfektion, och det kan också tas bort efter 4-6 timmar.
Hieff Trans™ levereras i steril flytande form. Vanligtvis, för 24-brunnars platttransfektion, cirka 1,5 μL varje gång, 1 mL Hieff Trans™ kan göra cirka 660 transfektioner; för en platta med 6 brunnar, cirka 6 μL varje gång, kan 1 mL Hieff Trans™ göra cirka 660 transfektioner. 160 transfektioner.
Särdrag
- Exceptionell effektivitet: Överlägsen transfektionsprestanda i ett brett spektrum av cellinjer, inklusive transient transfektion och stabil transfektion
- Bred anpassningsförmåga: Utmärkt transfektionseffektivitet i en mängd olika cellinjer och höga nivåer av rekombinant proteinuttryck
- Låg toxicitet: De olika cellernas aktiviteter nästan opåverkade av transfektionsreagenser från YEASEN
- Enkel användning: Bevisad effekt i närvaro av serum – eliminerar behovet av att byta media efter transfektion
- Kostnadseffektiv: Konkurrenskraftig transfektionseffekt med mer överkomliga priser
Ansökan
- Celltransfektion
Specifikation
| Form | Flytande |
| Serum kompatibel | Ja |
| Celltyp | Etablerade cellinjer |
| Provtyp | Plasmid DNA, syntetisk siRNA |
| Transfektionsteknik | Lipid-baserad transfektion |
Komponenter
| Komponentnr. | Namn | 40802ES02 | 40802ES03 | 40802ES08 |
| 40802 | Hieff Trans™ Liposomal transfektionsreagens | 0,5 ml | 1 ml | 5×1 ml |
Frakt och förvaring
Produkten levereras med isförpackningar och kan förvaras vid 2-8ºC i ett år. Frys inte!
Siffror

Citerat från: Communications Biology volym 5, Artikelnummer: 1296 (2022)
- Demonstration av transfektionseffekt
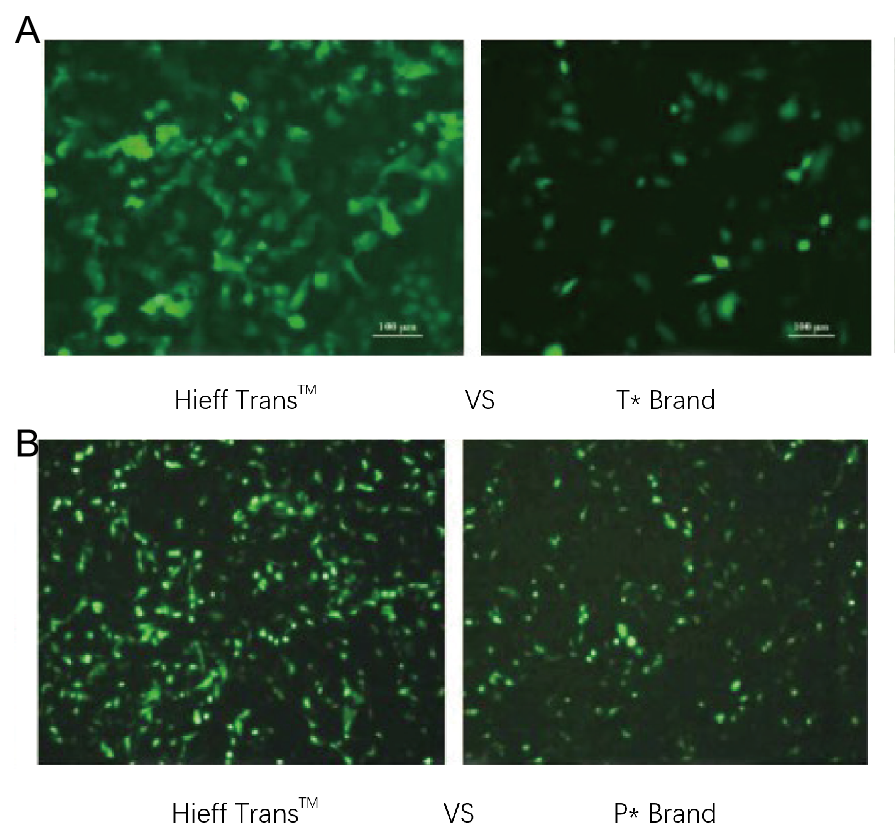
Figur 1. Hieff Trans™ liposomtransfektionsreagens överträffar transfektionsreagenset från det konkurrerande varumärket (T*-märket i figur 1a, P*-märket i figur 1b).
Varje reagens användes för att transfektera målcellinjen (Hela-cellinjen i figur 1a, DF-1-cellinjen i figur 1b) i ett format med 96 brunnar. GFP-uttryck analyserades 48 timmar efter transfektion. Hieff Trans™ Liposome Transfection Reagent gav högre GFP-transfektionseffektivitet än de konkurrerande varumärkesprodukterna.
- Validerade cellinjer
| Produkt Namn | Hieff TransTMLiposomal Transfektion Reagens | |||||
| Validerad Cell Rader | 293T | Calu 1 | HEK293 | HO1980 | N2A | TS |
| 293F | CHO-K1 | HEK293T | HUVEC | NCI-H1975 | U-87 | |
| 293FT | COS-7 | Hela | MCF10A | NIH-3T3 | Vero | |
| 3T3 | DF-1 | Hep2C | MCF-7 | Neuro-2a | WEHI | |
| A549 | H1299 | Hep3B | MDA-MB-231 | PC-12 | WRL-68 | |
| BV-2 | H520 | Hepa1-6 | MDA-MB-231-LM2-4175 | Raw264.7 | ||
| B50 | HaCaT | HepG2 | MDCK | SGC-7901 | ||
| C2C12 | HCT116 | HK2 | MEF | T47D | … | |
Figur 2. Lista över cellinjer som framgångsrikt transfekterats med Hieff Trans™ Liposomal transfektionsreagens (Under kontinuerlig uppdatering).

Citerad från "Optogenetic control of RNA function and metabolism using engineered light-switchable RNA-binding proteins. Nat Biotechnol. 2022 Jan 3. doi: 10.1038/s41587-021-01112-1"

Citerad från "Structural insight into BRCA1-BARD1 complex recruitment to damaged chromatin. Mol Cell. 2021 Jul 1;81(13):2765-2777.e6. doi: 10.1016/j.molcel.2021.05.010."
Citerad från "UBQLN2-HSP70-axeln reducerar poly-Gly-Ala-aggregat och lindrar beteendedefekter i C9ORF72-djurmodellen. Neuron. 2021 Jun 16;109(12):1949-1962.e6. doi: 10.1016/j.2021neur.."
Hieff Trans™ Liposomal transfektionsreagens FAQ
(1) F: Kan serum vara närvarande vid beredning av nukleinsyratransfektionsreagenskomplex?
S: Närvaron av serum kommer att påverka bildandet av liposomer. Det rekommenderas att använda serumfritt medium (vanligtvis MEM-medium) vid beredning av nukleinsyratransfektionsreagenskomplex.
(2) F: Kan transfektionsreagenset frysas?
S: Nej. Detta reagens måste förvaras vid 2-8 ℃, och försiktighet bör iakttas för att undvika att öppna locket upprepade gånger under lång tid, eftersom långvarig öppning av locket kommer att orsaka liposomoxidation och påverka transfektionseffektiviteten.
(3) F: Vad ska jag vara uppmärksam på när jag använder Hieff Trans™ liposomnukleinsyratransfektionsreagens?
S: 1) Under transfektionsoperationen är det bättre att cellkonfluensen når 80%-95%, och den specifika pläteringsdensiteten bestäms i enlighet med cellernas situation;
2) Att använda högrent DNA hjälper till att erhålla högre transfektionseffektivitet;
3) DNA och transfektionsreagens måste spädas med serumfritt medium vid framställning av transfektionskomplex;
4) Antibiotika kan inte tillsättas till mediet under transfektion;
5) DNA-koncentrationen och mängden katjonisk liposomreagens bör optimeras för den första användningen för att uppnå maximal transfektionseffektivitet. Förhållandet mellan DNA och transfektionsreagens är rekommenderas generellt att vara 1:2-1:3.
(4) F: Behöver den avslutas efter transfektion?
A: Inget behov. Liposomkomplex är stabila i 6 timmar. Om cellmediet inte byts före transfektion, för att säkerställa de näringsämnen som krävs för normal celltillväxt, är det nödvändigt att byta till ett nytt medium efter 4 till 6 timmar. Men om mediet har bytts före transfektion är det inte nödvändigt att byta medium efter liposomtransfektion.
(5) F: Vad ska jag vara uppmärksam på om jag vill förbättra transfektionseffektiviteten?
A: a: Tätheten av celler vid tidpunkten för transfektion är 90%-95%.
b: Under transfektion, använd MEM-serumfritt medium för nukleinsyra- och liposomspädningar.
c: Mediet kan bytas 4-6 timmar efter transfektion.
(6) F: Kan samtransfektion av DNA och siRNA utföras? Hur är effekten?
S: Samtransfektion kan utföras, men det rekommenderas att utföra separat transfektion, och DNA-transfektion bör utföras 6 timmar efter siRNA. Om de opereras tillsammans kommer siRNA-transfektionseffektiviteten att bli sämre.
(7) F: Kan transfektionsreagenset användas för lentiviral förpackningstransfektion?
S: Lentiviral förpackning är möjlig, men effektiviteten av lentiviral förpackning är inte nödvändigtvis relaterad till effektiviteten av transfektion, utan också relaterad till valet av förpackningsplasmider och förhållandet mellan plasmider.
(8) F: Kan Hieff Trans™ liposomnukleinsyratransfektionsreagens användas för transfektion av suspensionsceller?
S: Hieff Trans™ liposomnukleinsyratransfektionsreagens kan användas för suspensionscelltransfektion, se protokoll för detaljer. Dessutom har vi även lanserat ett transfektionsreagens specifikt för suspensionsceller (kat. nr. 40805, liposomnukleinsyratransfektionsreagens för suspensionsceller).
[1] Liu R, Yang J, Yao J, Zhao Z, He W, Su N, Zhang Z, Zhang C, Zhang Z, Cai H, Zhu L, Zhao Y, Quan S, Chen X, Yang Y. Optogenetisk kontroll av RNA-funktion och metabolism med hjälp av konstruerade ljusomkopplingsbara RNA-bindande proteiner. Nat Biotechnol. 2022 Jan 3. doi: 10.1038/s41587-021-01112-1. Epub före tryck. PMID: 34980910. (IF:54.908)
[2] Zhou J, Chen P, Wang H, Liu H, Li Y, Zhang Y, Wu Y, Paek C, Sun Z, Lei J, Yin L. Cas12a-varianter designade för lägre genomomfattande effekt utanför målet genom strikt PAM-igenkänning. Mol Ther. 5 januari 2022;30(1):244-255. doi: 10.1016/j.ymthe.2021.10.010. Epub 2021 20 oktober. PMID: 34687846; PMCID: PMC8753454. (IF:11.454)
[3] Chen S, Cao X, Zhang J, Wu W, Zhang B, Zhao F. circVAMP3 driver CAPRIN1 fasseparation och hämmar hepatocellulärt karcinom genom att undertrycka c-Myc Translation. Adv Sci (Weinh).2022 Mar;9(8):e2103817. doi: 10.1002/advs.202103817. Epub 2022 24 jan. PMID: 35072355; PMCID: PMC8922094. (IF:16.808)
[4] Zhang Y, Yu X, Sun R, Min J, Tang X, Lin Z, Xie S, Li X, Lu S, Tian Z, Gu C, Teng L, Yang Y. Splitsningsfaktor arginin/serinrik 8 främjar multipelt myelom malignitet och benskada genom alternativ splitsning av CACYBP och exosombaserad cellulär kommunikation. Clin Transl Med. 2022 feb;12(2):e684. doi: 10.1002/ctm2.684. PMID: 35184390. (IF:11.492)
[5] Tang X, Deng Z, Ding P, Qiang W, Lu Y, Gao S, Hu Y, Yang Y, Du J, Gu C. Ett nytt protein som kodas av circHNRNPU främjar multipelt myelomprogression genom att reglera benmärgens mikromiljö och alternativ splitsning. J Exp Clin Cancer Res. 2022 mars 8;41(1):85. doi: 10.1186/s13046-022-02276-7. PMID: 35260179. (IF:11.161)
[6] Hua Z, Wei R, Guo M, Lin Z, Yu X, Li X, Gu C, Yang Y. YTHDF2 främjar cellproliferation av multipelt myelom via STAT5A/MAP2K2/p-ERK-axeln. Onkogen. 2022 Mar;41(10):1482-1491. doi: 10.1038/s41388-022-02191-3. Epub 2022 24 januari. PMID: 35075244. (IF:9.867)
[7] Liang Y, Lu Q, Li W, Zhang D, Zhang F, Zou Q, Chen L, Tong Y, Liu M, Wang S, Li W, Ren X, Xu P, Yang Z, Dong S, Zhang B, Huang Y, Li D, Wang H, Yu W. Reaktivering av tumörsuppressor vid bröstcancer genom förstärkarväxling genom NamiRNA-nätverk. Nucleic Acids Res. 2021 sep 7;49(15):8556-8572. doi: 10.1093/nar/gkab626. PMID: 34329471; PMCID: PMC8421228. (IF:16,9)
[8] Dai L, Dai Y, Han J, Huang Y, Wang L, Huang J, Zhou Z. Strukturell insikt i BRCA1-BARD1 komplex rekrytering till skadat kromatin. Mol Cell. 2021 juli 1;81(13):2765-2777.e6. doi: 10.1016/j.molcel.2021.05.010. Epub 2021 7 juni. PMID: 34102105. (IF:17.97)
[9] Zhang K, Wang A, Zhong K, Qi S, Wei C, Shu X, Tu WY, Xu W, Xia C, Xiao Y, Chen A, Bai L, Zhang J, Luo B, Wang W, Shen C. UBQLN2-HSP70-axeln reducerar poly-Gly-Ala aggregerar i djurmodellen C9 och lindrar7 beteendedefekter ORF2. Neuron. 2021 Jun 16;109(12):1949-1962.e6. doi: 10.1016/j.neuron.2021.04.023. Epub 2021 14 maj. PMID: 33991504. (IF:17.17)
[10] Liang Y, Lu Q, Li W, Zhang D, Zhang F, Zou Q, Chen L, Tong Y, Liu M, Wang S, Li W, Ren X, Xu P, Yang Z, Dong S, Zhang B, Huang Y, Li D, Wang H, Yu W. Reaktivering av tumörsuppressor vid bröstcancer genom förstärkarnätverksbyte genom NamiRNA-nätverk. Nucleic Acids Res. 2021 sep 7;49(15):8556-8572. doi: 10.1093/nar/gkab626. PMID: 34329471; PMCID: PMC8421228. (IF:16,9)
[11] Li T, Chen X, Qian Y, Shao J, Li X, Liu S, Zhu L, Zhao Y, Ye H, Yang Y. En syntetisk BRET-baserad optogenetisk enhet för pulserande transgenuttryck som möjliggör glukoshomeostas hos möss. Nat Commun. 2021 jan 27;12(1):615. doi: 10.1038/s41467-021-20913-1. PMID: 33504786; PMCID: PMC7840992. (IF:14,92)
[12] Pan Y, He X, Li C, Li Y, Li W, Zhang H, Wang Y, Zhou G, Yang J, Li J, Qu J, Wang H, Gao Z, Shen Y, Li T, Hu H, Ma H. Neuronal aktivitet rekryterar CRTC1/CREB-axeln för att driva transkriptionsberoende autofagi LTD. Cell Rep. 2021 Jul 20;36(3):109398. doi: 10.1016/j.celrep.2021.109398. PMID: 34289350. (IF:9.42)
[13] Liu H, Xing R, Ou Z, Zhao J, Hong G, Zhao TJ, Han Y, Chen Y. G-proteinkopplad receptor GPR17 hämmar gliomutveckling genom att öka polycomb-repressivt komplex 1-medierad ROS-produktion. Celldöd Dis. 2021 juni 12;12(6):610. doi: 10.1038/s41419-021-03897-0. PMID: 34120140; PMCID: PMC8197764. (IF:8.463)
[14] Fan Y, Wang J, Jin W, Sun Y, Xu Y, Wang Y, Liang X, Su D. CircNR3C2 främjar HRD1-medierad tumörsuppressiv effekt via sponging miR-513a-3p vid trippelnegativ bröstcancer. Mol Cancer. 2021 februari 2;20(1):25. doi: 10.1186/s12943-021-01321-x.PMID: 33530981; PMCID: PMC7851937. (IF:27.403)
[15] Gu C, Wang Y, Zhang L, Qiao L, Sun S, Shao M, Tang X, Ding P, Tang C, Cao Y, Zhou Y, Guo M, Wei R, Li N, Xiao Y, Duan J, Yang Y. AHSA1 är ett lovande terapeutiskt mål för cellulär multipelproliferation och proteasomhämmareresistens. J Exp Clin Cancer Res. 2022 jan 6;41(1):11. doi: 10.1186/s13046-021-02220-1. PMID: 34991674; PMCID: PMC8734095. (IF:11.161)
[16] Luo Q, Wu X, Zhao P, Nan Y, Chang W, Zhu X, Su D, Liu Z. OTUD1 Aktiverar kaspasoberoende och kaspasberoende apoptos genom att främja AIF-kärntranslokation och MCL1-nedbrytning. Adv Sci (Weinh). 8 februari 2021; 8(8):2002874. doi: 10.1002/advs.202002874. PMID: 33898171; PMCID: PMC8061361. (IF:15,84)
[17] Luo Q, Wu X, Nan Y, Chang W, Zhao P, Zhang Y, Su D, Liu Z. TRIM32/USP11 balanserar ARID1A-stabilitet och det onkogena/tumörsuppressiva tillståndet för skivepitelcancer. Cell Rep. 2020 Jan 7;30(1):98-111.e5. doi: 10.1016/j.celrep.2019.12.017. PMID: 31914402. (IF:9.42)
[18] Sun X, Peng X, Cao Y, Zhou Y, Sun Y. ADNP främjar neural differentiering genom att modulera Wnt/β-catenin-signalering. Nat Commun. 12 juni 2020;11(1):2984. doi: 10.1038/s41467-020-16799-0. PMID: 32533114; PMCID: PMC7293280. (IF:14.911)
[19] Yang X, Wang H, Xie E, Tang B, Mu Q, Song Z, Chen J, Wang F, Min J. Omkoppling av ERBB3- och ERK-signalering ger resistens mot FGFR1-hämning i mag-tarmcancer med en ERBB3-E928G-mutation. Proteincell. 2020 Dec;11(12):915-920. doi: 10.1007/s13238-020-00749-z. PMID: 32632529; PMCID: PMC7719122. (IF:14.872)
[20] Chen, T., Chen, Y., Chen, H. et al. Dubbel-enzym-driven obegränsad DNA-gående nanomaskin för intracellulär avbildning av lågt uttryckt mikroRNA. Nano Res. 12, 1055–1060 (2019). https://doi.org/10.1007/s12274-019-2344-5 (IF:8.21)
[21] Zhang X, Qi Z, Yin H, Yang G. Interaktion mellan p53 och Ras-signalering styr cisplatinresistens via HDAC4- och HIF-1α-medierad reglering av apoptos och autofagi. Teranostik. 2019 jan 30;9(4):1096-1114. doi: 10.7150/thno.29673. PMID: 30867818; PMCID: PMC6401400. (IF:8.12)
[22] Zou Y, Wang A, Shi M, Chen X, Liu R, Li T, Zhang C, Zhang Z, Zhu L, Ju Z, Loscalzo J, Yang Y, Zhao Y. Analys av redoxlandskap och dynamik i levande celler och in vivo med hjälp av genetiskt kodade fluorescerande sensorer. Nat Protoc. 2018 okt;13(10):2362-2386. doi: 10.1038/s41596-018-0042-5. PMID: 30258175; PMCID: PMC6714056. (IF:13.49)
[23] Zhang K, Zhao X, Chen X, Wei Y, Du W, Wang Y, Liu L, Zhao W, Han Z, Kong D, Zhao Q, Guo Z, Han Z, Liu N, Ma F, Li Z. Förbättrade terapeutiska effekter av mesenkymala stamcellshärledda exosomer med en injicerbar hydrokemi. ACS Appl Mater-gränssnitt. 2018 sep 12;10(36):30081-30091. doi: 10.1021/acsami.8b08449. Epub 2018 29 augusti. PMID: 30118197. (IF:8.09)
[24] Hao H, Hu S, Chen H, Bu D, Zhu L, Xu C, Chu F, Huo X, Tang Y, Sun X, Ding BS, Liu DP, Hu S, Wang M. Förlust av endotelial CXCR7 försämrar vaskulär homeostas och hjärtremodellering efter hjärtinfarkt: Upptäcker hjärtinfarkt: implikationer. Omlopp. 2017 mars 28;135(13):1253-1264. doi: 10.1161/CIRCULATIONAHA.116.023027. Epub 2017 2 februari. PMID: 28154007. (IF:18.881)
Betalning och säkerhet
Din betalningsinformation behandlas säkert. Vi lagrar inte kreditkortsuppgifter och har inte heller tillgång till din kreditkortsinformation.
Förfrågan
Du kanske också gillar
Vanliga frågor
Produkten är endast avsedd för forskningsändamål och är inte avsedd för terapeutisk eller diagnostisk användning hos människor eller djur. Produkter och innehåll skyddas av patent, varumärken och upphovsrätt som ägs av Yeasen Biotechnology. Varumärkessymboler anger ursprungsland, inte nödvändigtvis registrering i alla regioner.
Vissa applikationer kan kräva ytterligare immateriella rättigheter från tredje part.
Yeasen är dedikerad till etisk vetenskap, och anser att vår forskning bör behandla kritiska frågor samtidigt som den säkerställer säkerhet och etiska standarder.