





وصف
يتم تحضير حبيبات اختيار الحمض النووي Hieff NGS™ بناءً على مبدأ SPRI (التثبيت العكسي في الطور الصلب) وهي قابلة للتطبيق لتنقية الحمض النووي واختيار الحجم أثناء تحضير مكتبات التسلسل الجيني للجيل التالي (NGS). حبيبات اختيار الحمض النووي Hieff NGS™ متوافقة مع العديد من مجموعات تحضير مكتبات الحمض النووي والحمض النووي الريبي وهي بديل جيد لـ حبات AMPure.
عناصر
| المكونات رقم | اسم | 12601ES08 | 12601ES56 | 12601ES75 |
| 12601 | خرز اختيار الحمض النووي Hieff NGS™ | 5 مل | 60 مل | 450 مل |
تحديد
| خط المنتج | تنظيف واختيار حبات الحمض النووي |
| المواد الأولية | الحمض النووي |
| التوافق | الحمض النووي |
| تكنولوجيا العزل | خرزة مغناطيسية |
| نوع المنتج النهائي | الحمض النووي |
| للاستخدام مع (التطبيق) | تنقية الحمض النووي، اختيار حجم الحمض النووي |
الشحن والتخزين
يتم شحن الخرز مع أكياس الثلج ويمكن تخزينها في درجة حرارة تتراوح بين 2 درجة مئوية و8 درجات مئوية لمدة عام واحد.
تعليمات
- 1. التحضير
قم بموازنة حبات الاختيار في درجة حرارة الغرفة لمدة 30 دقيقة على الأقل قبل الاستخدام.
- 2. اختيار حجم الحمض النووي
يظهر تدفق عملية اختيار الحجم في الشكل 1 والبروتوكول على النحو التالي.
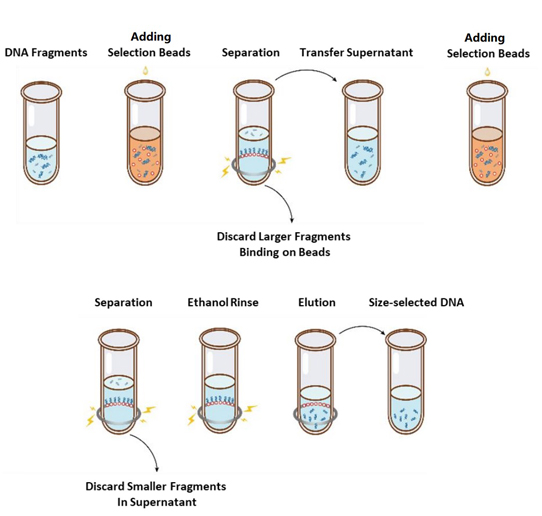
الشكل 1. مخطط انسيابي لاختيار حجم الحمض النووي
2.1 قم بخلط الخرز جيدًا عن طريق التحريك أو التقطير لأعلى ولأسفل في كل مرة قبل الاستخدام.
2.2 أضف الجولة الأولى من خرزات الاختيار إلى العينة (راجع الجدول 1). اخلط جيدًا عن طريق المزج بالدوامة أو التقطير لأعلى ولأسفل 10 مرات على الأقل.
2.3 احضنها في درجة حرارة الغرفة لمدة 5 دقائق.
2.4 قم بتدوير الأنبوب لأسفل لفترة وجيزة ثم ضعه على حامل مغناطيسي. عندما يصبح المحلول صافيًا (حوالي 5 دقائق)، انقل السائل العلوي إلى أنبوب PCR جديد.
2.5 أضف الجولة الثانية من خرزات الاختيار إلى العينة من الخطوة 2.4 وفقًا للجدول 1. اخلط جيدًا عن طريق المزج بالدوامة أو التقطير لأعلى ولأسفل 10 مرات على الأقل.
2.6 احضنها في درجة حرارة الغرفة لمدة 5 دقائق.
2.7 قم بتدوير الأنبوب لأسفل لفترة وجيزة ثم ضعه على حامل مغناطيسي. عندما يصبح المحلول صافيًا (حوالي 5 دقائق)، قم بشفط السائل العلوي وتخلص منه.
2.8 ضع الأنبوب في الحامل المغناطيسي وأضف 200 ميكرولتر من الإيثانول الطازج بنسبة 80% دون إزعاج الخرز، ثم احتضنه في درجة حرارة الغرفة لمدة 30 ثانية. قم بتصفية الإيثانول وتخلص منه.
2.9 كرر الخطوة 2.8 مرة واحدة لإجمالي غسلتين.
2.10 قم بإزالة الإيثانول المتبقي باستخدام أطراف ماصة سعة 10 ميكرولتر. احتفظ بالأنبوب في الحامل المغناطيسي، ثم قم بتجفيف حبات الاختيار في الهواء مع فتح الغطاء حتى تظهر الشقوق (حوالي 5 دقائق).
ملحوظة: لا تجفف حبات التحديد بشكل مفرط. قد يؤدي هذا إلى انخفاض معدل استرداد الحمض النووي المستهدف.
2.11 قم بإزالة الأنبوب من الحامل المغناطيسي. أضف كمية مناسبة من ddH2O (≥20 µL) واخلط جيدًا عن طريق المزج بالدوامة أو التقطير لأعلى ولأسفل 10 مرات على الأقل.
2.12. احضنيه في درجة حرارة الغرفة لمدة 5 دقائق.
قم بتدوير الأنبوب لفترة وجيزة ثم ضعه على الحامل المغناطيسي. عندما يصبح المحلول صافيًا (حوالي 5 دقائق)، انقل 20 ميكرولترًا من السائل العلوي إلى أنبوب جديد.
- 3. الشروط الموصى بها لاختيار حجم الحمض النووي
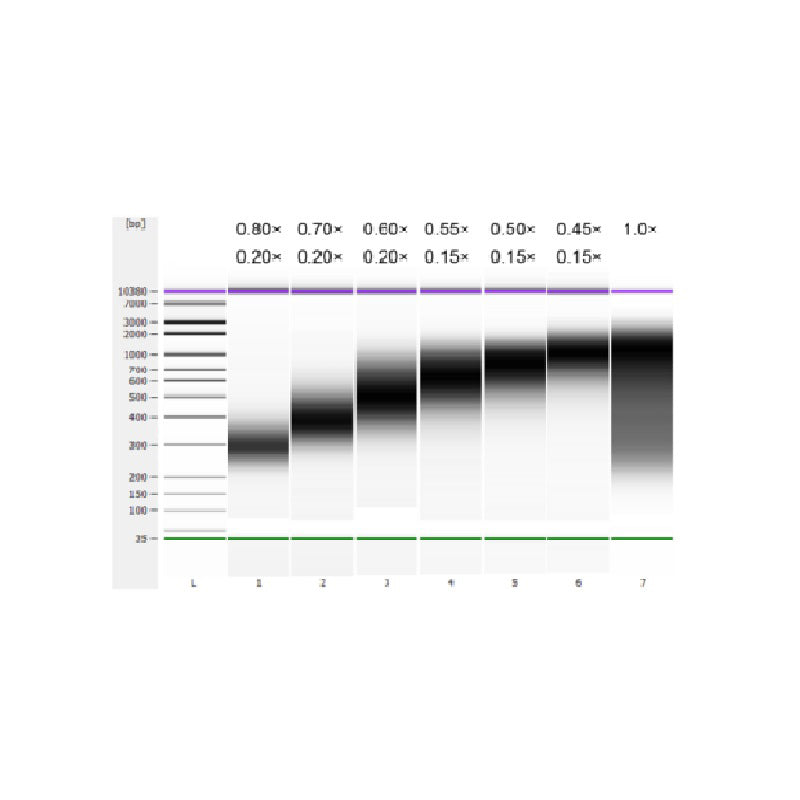
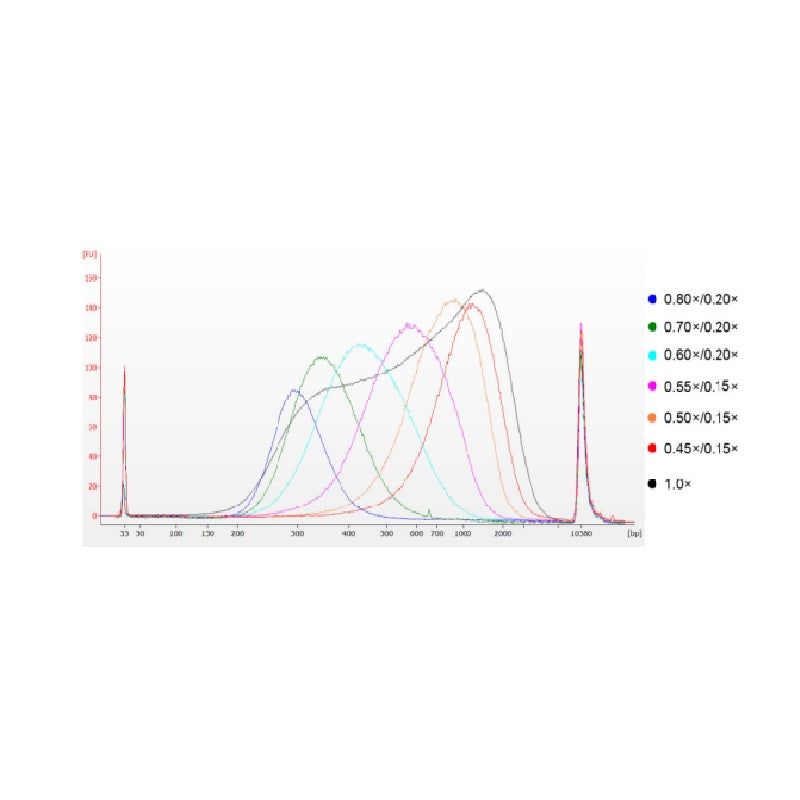
تم تجزئة الحمض النووي لغدة الزعترية العجلية باستخدام الموجات فوق الصوتية لإعداد قطعة بطول 100-1000 زوج أساسي، وتم إجراء جولتين لاختيار الحجم وفقًا للجدول 1. وتم تحليل النتائج باستخدام جهاز Agilent 2100 Bioanalyzer (الشكل 2).
الجدول 1. الحالة الموصى بها لاختيار حجم الحمض النووي
| طول قطعة الحمض النووي | 250-350 نقطة أساس | 320-420 زوجًا أساسيًا | 450-550 زوج أساسي | 550-700 زوج أساسي | 700-900 زوج أساسي | 800-1000 زوج أساسي |
| نسبة الخرز: DNA للجولة الأولى | 0.80× | 0.70× | 0.60× | 0.55× | 0.50× | 0.45× |
| نسبة الخرز: DNA للصف الثاني دائري | 0.20× | 0.20× | 0.20× | 0.15× | 0.15× | 0.15× |
ملاحظة: يشير "×" في الجدول إلى حجم عينة الحمض النووي. على سبيل المثال، إذا كان طول إدراج المكتبة 250 زوجًا قاعديًا وكان حجم عينة الحمض النووي 100 ميكرولتر، فإن حجم الخرز المغناطيسي المستخدم في الجولة الأولى من الفرز هو 0.80×100 ميكرولتر = 80 ميكرولتر؛ وحجم الخرز المغناطيسي المستخدم في الجولة الثانية من الفرز هو 0.20×100 ميكرولتر = 20 ميكرولتر.
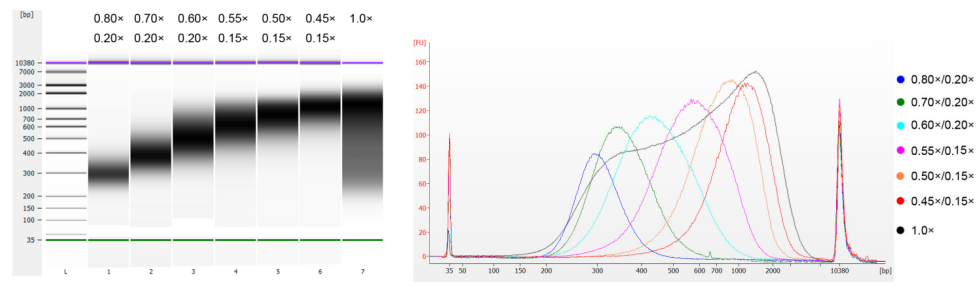
الشكل 2. مخطط كهربية الحمض النووي عالي الحساسية من Agilent 2100
ملحوظات:
1. من أجل سلامتك وصحتك، يرجى ارتداء المعاطف المخبرية والقفازات التي تستخدم لمرة واحدة أثناء العمل.

مقتبس من "التكامل المحدد للتسلسل بواسطة عائلة 1 من كاسبوزاز من Candidatus Nitrosopumilus koreensis AR1. Nucleic Acids Res. 2021؛49(17):9938-9952. doi:10.1093/nar/gkab725"

مقتبس من "العدوى الحديثة ببكتيريا Wolbachia تغير المجتمعات الميكروبية في مجموعات Laodelphax striatellus البرية. Microbiome. 2020؛8(1):104. نُشر في 2 يوليو 2020. doi:10.1186/s40168-020-00878-x"
[1] Wang X، Yuan Q، Zhang W، وآخرون. التكامل المحدد للتسلسل بواسطة عائلة 1 من كاسبوزاز من Candidatus Nitrosopumilus koreensis AR1. Nucleic Acids Res. 2021؛49(17):9938-9952. doi:10.1093/nar/gkab725(IF:16.971)
[2] Duan XZ، Sun JT، Wang LT، وآخرون. العدوى الحديثة ببكتيريا Wolbachia تغير المجتمعات الميكروبية في مجموعات Laodelphax striatellus البرية. Microbiome. 2020؛8(1):104. نُشر في 2 يوليو 2020. doi:10.1186/s40168-020-00878-x(IF:11.607)
[3] سونغ ب، المطرفي إي، سانغ ف، وآخرون.إدارة الرواسب المعالجة بفينتون باستخدام الفحم الحيوي وسماد روث الأغنام: التأثيرات على الخصائص التطورية للمجتمع البكتيري. مجلة إدارة البيئة. 2022؛ 316: 115218. doi: 10.1016/j.jenvman.2022.115218(IF: 6.789)
[4] Huang C, Mei Q, Lou L, et al. Ulcerative Colitis in Response to Fecal Microbiota Transplantation via Modulation of Gut Microbiota and Th17/Treg Cell Balance. Cells. 2022;11(11):1851. نُشر في 5 يونيو 2022. doi:10.3390/cells11111851(IF:6.600)
[5] Ghosh S, Yang X, Wang L, Zhang C, Zhao L. تعمل التغذية بالبريبايوتيك في المرحلة النشطة على تغيير ميكروبات الأمعاء، وتحفز تخفيفًا مستقلًا عن الوزن لتدهن الكبد والكوليسترول في المصل لدى الفئران التي تتغذى على نظام غذائي غني بالدهون. مجلة Comput Struct Biotechnol J. 2020؛ 19: 448-458. نُشر في 24 ديسمبر 2020. doi:10.1016/j.csbj.2020.12.011(IF:6.018)
[6] Gao X، Yu B، Yu J، وآخرون. ملف تعريف النمو لهضم الكربوهيدرات الغذائية في الخنازير. Front Microbiol. 2022؛ 13: 896660. نُشر في 29 أبريل 2022. doi:10.3389/fmicb.2022.896660(IF:5.640)
[7] Li P, Zhang Y, Yan F, Zhou X. خصائص البكتيريا، vB_Kox_ZX8، المعزولة من Klebsiella oxytoca السريرية وتأثيرها العلاجي على بكتيريا الدم لدى الفئران. Front Microbiol. 2021؛12:763136. نُشر في 3 ديسمبر 2021. doi:10.3389/fmicb.2021.763136(IF:5.640)
[8] Lin Z، Luo P، Huang D، Wu Y، Li F، Liu H. استراتيجية متعددة الأوميكس لتحليل سمية الأكريلاميد في نموذج Saccharomyces cerevisiae. Chem Biol Interact. 2021؛349:109682. doi:10.1016/j.cbi.2021.109682(IF:5.194)
[9] صن إكس، إل في دبليو، وانغ واي، وآخرون. يلعب جين Mrgprb2 دورًا في التفاعلات التأقية الناجمة عن حقن Houttuynia cordata. ي اثنوفارماكول. 2022;289:115053. دوى:10.1016/j.jep.2022.115053(IF:4.360)
[10] Ma H, Lai B, Zan C, Di X, Zhu X, Wang K. GLO1 يساهم في مقاومة الأدوية لـ Escherichia coli من خلال تحريض نوع PER من β-Lactamases ذات الطيف الممتد. Infect Drug Resist. 2022؛15:1573-1586. نُشر في 5 أبريل 2022. doi:10.2147/IDR.S358578(IF:4.003)
[11] Zhong Y, Zhao W, Tang Z, et al. Comparative transcriptomic analysis of the different developmental stage of ovary in red swamp crayfish Procambarus clarkii. BMC Genomics. 2021;22(1):199. نُشر في 21 مارس 2021. doi:10.1186/s12864-021-07537-x(IF:3.969)
[12] Lian C, Yang H, Lan J, et al. Comparative analysis of chloroplast genomes reveals phylogenetic relations and intraspecific variation in the medical plant Isodon rubescens. PLoS One. 2022;17(4):e0266546. Published 2022 Apr 6. doi:10.1371/journal.pone.0266546(IF:3.240)
[13] دياو جي، هوانغ جي، تشنغ إكس، وآخرون. البروستاجلاندين يلعب E2 دورًا مزدوجًا في تنظيم هجرة الخلايا الشجيرية. Int J Mol Med. 2021؛47(1):207-218. doi:10.3892/ijmm.2020.4801(IF:3.098)
[14] Bing XL، Zhao DS، Peng CW، Huang HJ، Hong XY. أوجه التشابه والاختلافات المكانية للمجتمعات البكتيرية والفطرية في مجموعات حشرات الأرز الحقلية (Hemiptera: Delphacidae). Insect Sci. 2020؛27(5):947-963. doi:10.1111/1744-7917.12782(IF:2.791)
[15] Li X, Zhou S, Zhang J, Zhou Z, Xiong Q. التغيرات الاتجاهية في مجتمع البكتيريا المعوية في يرقات ذبابة الجندي الأسود (Hermetia illucens). Animals (بازل). 2021؛ 11(12): 3475. نُشر في 6 ديسمبر 2021. doi:10.3390/ani11123475(IF:2.752)
[16] Yang J، Peng Y، Kong W. تحديد التسلسل الكامل لجينوم فيروس التوت الخفي 1. Arch Virol. 2022؛ 167 (2): 687-690. doi: 10.1007 / s00705-021-05350-1 (IF: 2.574)
[17] Chang Y، Xia X، Sui L، وآخرون. الاستعمار الداخلي للفطريات المسببة للأمراض الحشرية يزيد من مقاومة النبات للأمراض عن طريق تغيير مجتمع البكتيريا الداخلية. J Basic Microbiol. 2021؛ 61 (12): 1098-1112. doi: 10.1002/وظيفة.202100494(إذا:2.281)
[18] Ding CY, Ma YM, Li B, et al. Identification and Functional Analysis of Differentially Expressed Genes in Myzus persicae (Hemiptera: Aphididae) in Response to Trans-anethole. J Insect Sci. 2022;22(1):3. doi:10.1093/jisesa/ieab094(IF:1.857)
الدفع والأمن
تتم معالجة معلومات الدفع الخاصة بك بشكل آمن. لا نقوم بتخزين تفاصيل بطاقة الائتمان ولا يمكننا الوصول إلى معلومات بطاقة الائتمان الخاصة بك.
سؤال
قد تعجبك أيضًا
التعليمات
المنتج مخصص لأغراض البحث فقط وليس مخصصًا للاستخدام العلاجي أو التشخيصي لدى البشر أو الحيوانات. المنتجات والمحتوى محميان بموجب براءات الاختراع والعلامات التجارية وحقوق الطبع والنشر المملوكة لشركة Yeasen Biotechnology. تشير رموز العلامة التجارية إلى بلد المنشأ، وليس بالضرورة التسجيل في جميع المناطق.
قد تتطلب بعض التطبيقات حقوق الملكية الفكرية الإضافية لجهات خارجية.
تلتزم شركة Yeasen بالعلوم الأخلاقية، حيث تؤمن بأن أبحاثنا يجب أن تعالج الأسئلة الحرجة مع ضمان معايير السلامة والأخلاق.