

Beskrivelse
GMyc-PCR Mycoplasma Detection Kit bruger hovedsageligt PCR-metoden til at detektere Mycoplasma-infektion af forskellige biologiske materialer (såsom cellekultur, eksperimentelle dyresekreter, dyreserum osv.). Den kombinerer flere fordele: følsom, specifik, hurtig og kan påvises direkte med cellekultursupernatanter. Dette produkt detekterer mycoplasma i biologiske materialer såsom dyrkede celler ved PCR-metoden. De anvendte primere er designet i overensstemmelse med den konserverede region af 16S-23S rRNA-sekvensen af mycoplasma og amplificerer kun specifikt mycoplasma-DNA'et med høj detektionssensitivitet og specificitet. PCR-amplifikation og elektroforeseanalyse tager kun et par timer, og operationen er praktisk og enkel.
Cellekultur er et almindeligt eksperiment inden for biovidenskabelig forskning. I modsætning til andre almindeligt anvendte eksperimentelle metoder er cellekultur en dynamisk kontinuerlig proces, og celler reagerer ofte på manipulationsfejl eller kontaminanter, der ofte udviser unormale celletilstande eller medium udseende. Hvis det er forurenet af mycoplasma, har cellemorfologien ingen tydelig ændring, og det er let at blive overset. Det findes ofte først, når forureningen er meget alvorlig. Der kan være hundredvis af mycoplasmaer på den forurenede cellemembran, disse mycoplasmaer konkurrerer om næringsstoffer og frigiver giftige metabolitter, hvilket alvorligt påvirker de eksperimentelle resultater.
Undersøgelser har vist, at mindst 20 slags mycoplasma kan forurene celler, blandt hvilke de mest almindelige er: oral Mycoplasma (M. orale), Mycoplasma arginin (M. arginini), Mycoplasma hyorhinis (M. hyorhinis), Mycoplasma fermentum (M. fermentans), Mycoplasma hominis (M. hominis), Mycoplasma salivarius (M. salivarium), Mycoplasma pulmonal (M. pulmonis) og Mycoplasma pære (M. pirum). Mycoplasma-kontamineringen af dyrkede celler varierer fra 4 % til 92 %. Kilderne til kontaminering omfatter arbejdsmiljøet, operatøren selv (nogle mycoplasmaer er normal flora i den menneskelige krop), dyrkningsmedium, serum, cellekrydskontaminering, eksperimentelt udstyr og brugt kontaminering af det originale væv eller organ, hvorfra cellerne blev fremstillet.
At identificere den underliggende årsag til problemer under celledyrkning er en vanskelig og tidskrævende opgave, hvor pludselige ændringer bør mistænkes, og god testpraksis og regelmæssig testning for mycoplasma-kontamination er nødvendig. Der er mange metoder til påvisning af mycoplasma, såsom direkte dyrkning, DNA-fluorescensfarvning, ELISA og PCR-metoder.
Feature
- De anvendte primere blev designet i overensstemmelse med den konserverede region af Mycoplasma 16S-23S rRNA-sekvensen
- Kun amplificeret mycoplasma DNA specifikt
- Fremragende følsomhed og specificitet
- Mere end 20 mycoplasmaarter kunne påvises
Anvendelse
- Mycoplasma påvisning
Komponenter
| Komponenter nr. | Navn | 40601ES10 (10 analyser) | 40601ES20 (20 analyser) |
| 40601-A | GMyc-1st PCR-blanding | 250 µL | 2×250 µL |
| 40601-B | GMyc-2nd PCR Mix | 250 µL | 2×250 µL |
| 40601-C | Positiv kontrolskabelon N | 20 µL | 20 µL |
[Noter] 1. Når den ikke er i brug i lang tid, kan den opbevares frosset ved -85~-65℃.
2. PCR-reaktionen er ekstremt følsom.For at forhindre falske positiver tilføjes en positiv kontrol til sidst ved tilsætning af prøver.
Opbevaring
Dette produkt kan opbevares ved -25~-15 ℃ i 18 måneder. Hvis den ikke skal bruges i længere tid, bedes du holde den væk fra lys.
Figurer
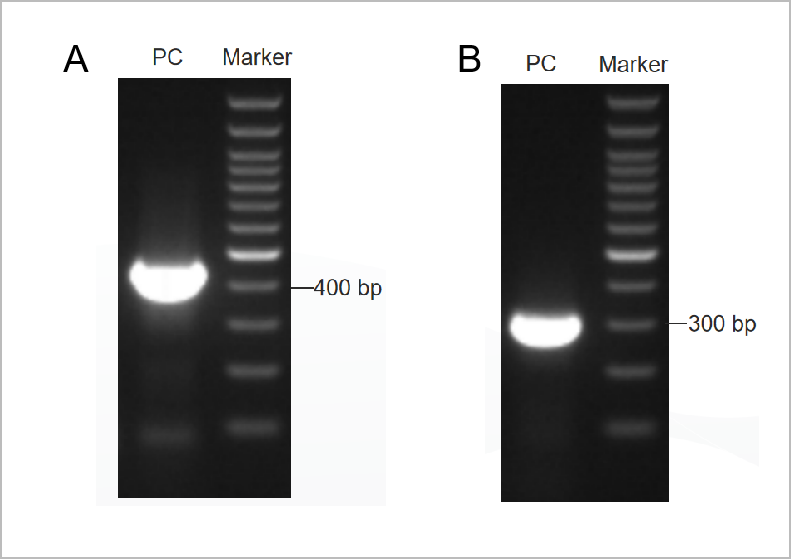
Figur 1. Elektroforetogramresultatet af PCR Mycoplama Test Kit.
Den første runde elektroforetogram blev vist i figur 1A, og den anden runde elektroforetogram blev vist i figur 1B. (M: 1 kb markør, 1-2: positive kontroller, 3: den negative kontrol)
[1] Rao XS, Cong XX, Gao XK, et al. AMPK-medieret phosphorylering øger auto-hæmningen af TBC1D17 for at fremme Rab5-afhængig glukoseoptagelse. Celledød er forskellig. 2021;28(12):3214-3234. doi:10.1038/s41418-021-00809-9(IF:15.828)
[2] Guo F, Li L, Li J, et al. Enkeltcellet multi-omics-sekventering af tidlige museembryoner og embryonale stamceller. Cell Res. 2017;27(8):967-988. doi:10.1038/cr.2017.82(IF:15.606)
[3] Hao Y, He B, Wu L, et al. Nuklear translokation af p85β fremmer tumorgenese af PIK3CA helical domæne mutant cancer. Nat Commun. 2022;13(1):1974. Udgivet 2022 13. april. doi:10.1038/s41467-022-29585-x(IF:14.919)
[4] Shu X, Liu M, Lu Z, et al. Genom-dækkende kortlægning afslører, at deoxyuridin er beriget i det humane centromeriske DNA. Nat Chem Biol. 2018;14(7):680-687. doi:10.1038/s41589-018-0065-9(IF:13.843)
[5] Li X, Xiong X, Wang K, et al. Transkriptom-dækkende kortlægning afslører reversibel og dynamisk N(1)-methyladenosin-methylom. Nat Chem Biol. 2016;12(5):311-316. doi:10.1038/nchembio.2040(IF:12.709)
[6] Sun L, Yang X, Huang X, et al. 2-Hydroxylering af fedtsyrer undertrykker kolorektal tumorgenese og metastase via YAP-transkriptionsaksen. Cancer Res. 2021;81(2):289-302. doi:10.1158/0008-5472.CAN-20-1517(IF:12.701)
[7] Sun Z, Zhang Z, Wang QQ, Liu JL. Kombineret inaktivering af CTPS1 og ATR er syntetisk dødelig for MYC-overudtrykkende kræftceller. Cancer Res. 2022;82(6):1013-1024. doi:10.1158/0008-5472.CAN-21-1707(IF:12.701)
[8] Song J, Zhuang Y, Zhu C, et al. Differentielle roller af human PUS10 i miRNA-behandling og tRNA-pseudouridylering. Nat Chem Biol. 2020;16(2):160-169. doi:10.1038/s41589-019-0420-5(IF:12.154)
[9] He B, Pan H, Zheng F, et al. Langt ikke-kodende RNA LINC00930 fremmer PFKFB3-medieret tumorglykolyse og celleproliferation i nasopharyngealt karcinom. J Exp Clin Cancer Res. 2022;41(1):77. Udgivet 2022 24. februar doi:10.1186/s13046-022-02282-9(IF:11.161)
[10] Tang B, Liu BH, Liu ZY, Luo MY, Shi XH, Pang DW. Quantum Dots med en kompakt amfifil Zwitterionisk belægning. ACS Appl Mater-grænseflader. 2022;14(24):28097-28104. doi:10.1021/acsami.2c04438(IF:9.229)
[11] Huang C, Zhang Z, Chen L, et al.Acetylering inden for de N- og C-terminale domæner af Src regulerer distinkte roller af STAT3-medieret tumorigenese. Cancer Res. 2018;78(11):2825-2838. doi:10.1158/0008-5472.CAN-17-2314(IF:9.130)
[12] Wu X, Yu M, Zhang Z, et al. DDB2 regulerer DNA-replikation gennem PCNA-uafhængig nedbrydning af CDT2. Cell Biosci. 2021;11(1):34. Udgivet 8. februar 2021. doi:10.1186/s13578-021-00540-5(IF:7.133)
[13] Wang J, Zhang Y, Liu X, Liu H. Optimering af adaptiv terapi baseret på nåbarheden til tumorresistent subpopulation. Kræft (Basel). 2021;13(21):5262. Udgivet 20. oktober 2021. doi:10.3390/cancers13215262(IF:6.639)
[14] Feng W, Liu R, Xie X, et al. SUMOylering af α-tubulin er en ny modifikation, der regulerer mikrotubulus dynamik. J Mol Cell Biol. 2021;13(2):91-103. doi:10.1093/jmcb/mjaa076(IF:6.216)
[15] Yu M, Hu X, Yan J, Wang Y, Lu F, Chang J. RIOK2-hæmmer NSC139021 udøver antitumoreffekter på glioblastom via inducering af Skp2-medieret cellecyklusarrest og apoptose. Biomedicin. 2021;9(9):1244. Udgivet 2021 17. sep. doi:10.3390/biomedicines9091244(IF:6.081)
[16] Ren S, Cai Y, Hu S, et al. Berberin udøver antitumoraktivitet i diffust storcellet B-celle lymfom ved at modulere c-myc/CD47-aksen. Biochem Pharmacol. 2021;188:114576. doi:10.1016/j.bcp.2021.114576(IF:5.858)
[17] Wen F, Sun X, Sun C, et al. TAGLN er nedreguleret af TRAF6-medieret proteasomal nedbrydning i prostatacancerceller. Mol Cancer Res. 2021;19(7):1113-1122. doi:10.1158/1541-7786.MCR-20-0513(IF:5.852)
[18] Tang B, Sun EZ, Zhang ZL, et al. Sphingomyelin-sekvestreret kolesteroldomæne rekrutterer Formin-bindende protein 17 til at forsnævre Clathrin-coatede gruber ved indtrængen af influenzavirus. J Virol. 2022;96(5):e0181321. doi:10.1128/JVI.01813-21(IF:5.103)
[19] Hu J, Ren W, Qiu W, et al. Generering af induceret pluripotent stamcellelinje (XDCMHi001-A) fra en ankyloserende spondylitis-patient med JAK2-mutation. Stamcelle Res. 2020;45:101788. doi:10.1016/j.scr.2020.101788(IF:4.495)
[20] Xiao S, Yao X, Ye J, Tian X, Yin Z, Zhou L. Epigenetisk modifikation letter prolinsyntase PYCR1 afvigende ekspression i mavekræft [publiceret online forud for tryk, 2022 30. maj]. Biochim Biophys Acta Gene Regul Mech. 2022;1865(6):194829. doi:10.1016/j.bbagrm.2022.194829(IF:4.490)
[21] Wang J, Zhang Y, Liu X, Liu H. Er den faste periodiske behandling effektiv for tumorsystemet uden fuldstændig information?. Cancer Manag Res. 2021;13:8915-8928. Udgivet 30. november 2021. doi:10.2147/CMAR.S339787(IF:3.989)
[22] Yang X, Ren S, Rehman ZU, et al. Molekylær karakterisering, ekspression og funktionel identifikation af TANK-bindende kinase 1 (TBK1) fra koen (Bos taurus) og geden (Capra hircus). Dev Comp Immunol. 2022;133:104444. doi:10.1016/j.dci.2022.104444(IF:3.636)
[23] Zheng D, Chang X, Liu Y, et al. 2-Methoxy-5((3,4,5-trimethosyphenyl)seleninyl)phenol reverserer EGF-induceret cellemigration og -invasion gennem nedregulering af MDM2 i brystkræftcellelinjer. Kræft Biol Ther. 2019;20(4):513-523. doi:10.1080/15384047.2018.1537578(IF:3.373)
[24] Xu F, Zhang S, Liu Z, et al. TEX9 og eIF3b synergerer funktionelt for at fremme progressionen af esophageal pladecellecarcinom. BMC Cancer. 2019;19(1):875. Udgivet 3. september 2019. doi:10.1186/s12885-019-6071-9(IF:2.933)
[25] Pan H, Sun L, Wang W, et al. Serum lang ikke-kodende RNA LOC553103 som ikke-specifik diagnostisk og prognostisk biomarkør for almindelige typer af human cancer. Clin Chim Acta. 2020;508:69-76. doi:10.1016/j.cca.2020.05.017(IF:2.615)
[26] Li N, Lin SM, Li Y, Sun J, Zhang L, Chen M.En induceret pluripotent stamcellelinje (GZHMCi004-A) afledt af et foster med heterozygot G380R-mutation i FGFR3-genet, der forårsager achondroplasi. Stamcelle Res. 2021;53:102322. doi:10.1016/j.scr.2021.102322(IF:2.020)
[27] Luo Q, Wei C, Long Y, et al. Generering af en ELTD1 knockout human embryonal stamcellelinje af iCRISPR/Cas9 systemet. Stamcelle Res. 2021;53:102350. doi:10.1016/j.scr.2021.102350(IF:2.020)
[28] Liu YQ, Ling TW, Wang HY, Yang YH, Song WJ, Wang TC. Generering af en integrationsfri induceret pluripotent stamcellelinje (LZUSHI001-A) fra en epileptisk patient med DGKG-mutation. Stamcelle Res. 2022;61:102768. doi:10.1016/j.scr.2022.102768(IF:2.020)
[29] Chen M, Lin SM, Li N, Li Y, Li Y, Zhang L. En induceret pluripotent stamcellelinje (GZHMCi003-A) afledt af et foster med exon 3 heterozygot deletion i RUNX2-genet, der forårsager cleidokraniel dysplasi. Stamcelle Res. 2021;51:102166. doi:10.1016/j.scr.2021.102166(IF:2.020)
[30] Xu Y, Wang X, Qiu T, et al. Generering af en induceret pluripotent stamcellelinje (FDCHI007-A) afledt af en patient med udviklingsmæssig og epileptisk encefalopati Type 31, der bærer heterozygot c.545C > En mutation i DNM1-genet. Stamcelle Res. 2022;60:102709. doi:10.1016/j.scr.2022.102709(IF:2.020)
[31] Fan T, He J, Wang Y, Yu J, Sun W. Generering af en induceret pluripotent stamcellelinje (FDCHi006-A) fra en 7-årig pige med central tidlig pubertet. Stamcelle Res. 2021;56:102542. doi:10.1016/j.scr.2021.102542(IF:2.020)
[32] Gong X, Zheng Z, Yang T, Zheng H, Xiao X, Jia N. Generering af en isogen genkorrigeret iPSC-linje (OGHFUi001-A-1) fra en type 1 tidlig infantil epileptisk encefalopati (EIEE1) patient med en hemizygot R330L-mutation i 330L-genet. Stamcelle Res. 2022;60:102693. doi:10.1016/j.scr.2022.102693(IF:2.020)
[33] Jia N, Gong X, Chen J, et al. Generering af en induceret pluripotent stamcellelinje (OGHFUi001-A) fra en type 1 tidlig infantil epileptisk encefalopati med ARX-mutation. Stamcelle Res. 2021;53:102367. doi:10.1016/j.scr.2021.102367(IF:2.020)
[34] Zhu W, Zhou Y, Wang Q, et al. Generering af en human induceret pluripotent stamcelle (iPSC) linje fra hudfibroblaster fra en patient, der bærer en E363Q-mutation i PSEN1-genet. Stamcelle Res. 2022;61:102769. doi:10.1016/j.scr.2022.102769(IF:2.020)
[35] Luo F, Long K, Li X, et al. Mangel på LRRC8A dæmper hypoxi-induceret nekrose i 3T3-L1-celler. Biosci Biotechnol Biochem. 2020;84(6):1139-1145. doi:10.1080/09168451.2020.1730689(IF:1.516)
Betaling og sikkerhed
Dine betalingsoplysninger behandles sikkert. Vi gemmer ikke kreditkortoplysninger og har heller ikke adgang til dine kreditkortoplysninger.
Forespørgsel
Du kan også lide
FAQ
Produktet er kun til forskningsformål og er ikke beregnet til terapeutisk eller diagnostisk brug hos mennesker eller dyr. Produkter og indhold er beskyttet af patenter, varemærker og ophavsrettigheder ejet af Yeasen Biotechnology. Varemærkesymboler angiver oprindelseslandet, ikke nødvendigvis registrering i alle regioner.
Visse applikationer kan kræve yderligere tredjeparts intellektuelle ejendomsrettigheder.
Yeasen er dedikeret til etisk videnskab og mener, at vores forskning bør adressere kritiske spørgsmål og samtidig sikre sikkerhed og etiske standarder.