


Beskrivelse
Hieff Trans™ Liposomal Transfection Reagent er et alsidigt liposomtransfektionsreagens, velegnet til DNA-, RNA- og oligonukleotidtransfektion, med høj transfektionseffektivitet for de fleste eukaryote celler. Dens unikke formel gør det muligt at tilføje det direkte til mediet, og tilstedeværelsen af serum påvirker ikke transfektionseffektiviteten, hvilket reducerer skaden på celler forårsaget af serumfjernelse. Det er ikke nødvendigt at fjerne nukleinsyre-Hieff Trans™-komplekset eller erstatte det med et frisk medium efter transfektion, og det kan også fjernes efter 4-6 timer.
Hieff Trans™ leveres i steril flydende form. Normalt, til 24-brønds pladetransfektion, ca. 1,5 μL hver gang, 1 mL Hieff Trans™ kan udføre omkring 660 transfektioner; for en 6-brønds plade, ca. 6 μL hver gang, kan 1 mL Hieff Trans™ udføre ca. 660 transfektioner. 160 transfektioner.
Feature
- Enestående effektivitet: Overlegen transfektionsydelse i en bred vifte af cellelinjer, herunder forbigående transfektion og stabil transfektion
- Bred tilpasningsevne: Fremragende transfektionseffektivitet i en række cellelinjer og høje niveauer af rekombinant proteinekspression
- Lav toksicitet: Aktiviteterne af de forskellige celler næsten upåvirket af transfektionsreagenser fra YEASEN
- Enkel betjening: Dokumenteret effektivitet i nærvær af serum - eliminerer behovet for at skifte medie efter transfektion
- Omkostningseffektiv: Konkurrencedygtig transfektionseffekt med mere overkommelige priser
Anvendelse
- Celletransfektion
Specifikation
| Form | Flydende |
| Serum kompatibel | Ja |
| Celletype | Etablerede cellelinjer |
| Prøvetype | Plasmid DNA, Syntetisk siRNA |
| Transfektionsteknik | Lipid-baseret transfektion |
Komponenter
| Komponenter nr. | Navn | 40802ES02 | 40802ES03 | 40802ES08 |
| 40802 | Hieff Trans™ Liposomal transfektionsreagens | 0,5 ml | 1 ml | 5×1 ml |
Forsendelse og opbevaring
Produktet sendes med isposer og kan opbevares ved 2-8ºC i et år. Må ikke fryses!
Figurer

Citeret fra: Communications Biology bind 5, artikelnummer: 1296 (2022)
- Demonstration af transfektionseffekt
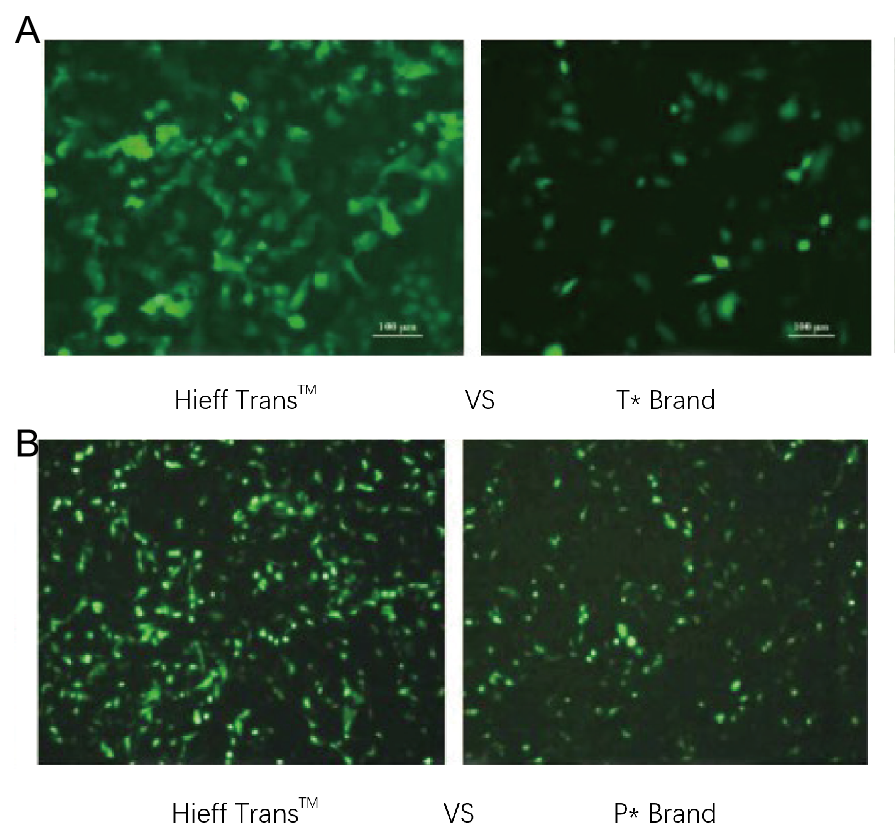
Figur 1. Hieff Trans™ liposomtransfektionsreagens overgår transfektionsreagenset fra det konkurrerende mærke (T*-mærket i figur 1a, P*-mærket i figur 1b).
Hvert reagens blev brugt til at transficere målcellelinjen (Hela-cellelinjen i figur 1a, DF-1-cellelinjen i figur 1b) i et 96-brønds format. GFP-ekspression blev analyseret 48 timer efter transfektion. Hieff Trans™ liposomtransfektionsreagens gav højere GFP-transfektionseffektivitet end de konkurrerende mærkeprodukter.
- Validerede cellelinjer
| Produkt Navn | Hieff TransTMLiposomal Transfektion Reagens | |||||
| Valideret Celle Linjer | 293T | Calu 1 | HEK293 | HO1980 | N2A | TS |
| 293F | CHO-K1 | HEK293T | HUVEC | NCI-H1975 | U-87 | |
| 293FT | COS-7 | Hela | MCF10A | NIH-3T3 | Vero | |
| 3T3 | DF-1 | Hep2C | MCF-7 | Neuro-2a | WEHI | |
| A549 | H1299 | Hep3B | MDA-MB-231 | PC-12 | WRL-68 | |
| BV-2 | H520 | Hepa1-6 | MDA-MB-231-LM2-4175 | Raw264.7 | ||
| B50 | HaCaT | HepG2 | MDCK | SGC-7901 | ||
| C2C12 | HCT116 | HK2 | MEF | T47D | … | |
Figur 2. Liste over cellelinjer, der er blevet transficeret med Hieff Trans™ Liposomal transfektionsreagens (Under løbende opdatering).

Citeret fra "Optogenetisk kontrol af RNA-funktion og metabolisme ved hjælp af konstruerede lys-switchable RNA-bindende proteiner. Nat Biotechnol. 2022 Jan 3. doi: 10.1038/s41587-021-01112-1"

Citeret fra "Strukturel indsigt i BRCA1-BARD1 kompleks rekruttering til beskadiget kromatin. Mol Cell. 2021 Jul 1;81(13):2765-2777.e6. doi: 10.1016/j.molcel.2021.05.010."
Citeret fra "UBQLN2-HSP70-aksen reducerer poly-Gly-Ala-aggregater og lindrer adfærdsdefekter i C9ORF72-dyremodellen. Neuron. 2021 Jun 16;109(12):1949-1962.e6. doi: 10.1016/j.2021neur.."
Hieff Trans™ Liposomal transfektionsreagens FAQ
(1) Spørgsmål: Kan serum være til stede ved fremstilling af nukleinsyretransfektionsreagenskompleks?
A: Tilstedeværelsen af serum vil påvirke dannelsen af liposomer. Det anbefales at bruge serumfrit medium (normalt MEM-medium) ved fremstilling af nukleinsyretransfektionsreagenskomplekser.
(2) Sp.: Kan transfektionsreagenset fryses?
A: Nej. Dette reagens skal opbevares ved 2-8 ℃, og man bør sørge for at undgå gentagne gange at åbne hætten i lang tid, da langvarig åbning af hætten vil forårsage liposomoxidation og påvirke transfektionseffektiviteten.
(3) Sp.: Hvad skal jeg være opmærksom på, når jeg bruger Hieff Trans™ liposomnukleinsyretransfektionsreagens?
A: 1) Under transfektionsoperationen er det bedre, at cellekonfluensen når 80%-95%, og den specifikke pletteringstæthed bestemmes i henhold til cellernes situation;
2) Brug af DNA med høj renhed hjælper med at opnå højere transfektionseffektivitet;
3) DNA og transfektionsreagenser skal fortyndes med serumfrit medium ved fremstilling af transfektionskomplekser;
4) Antibiotika kan ikke tilsættes til mediet under transfektion;
5) DNA-koncentrationen og mængden af kationisk liposomreagens bør optimeres til den første brug for at opnå den maksimale transfektionseffektivitet. Forholdet mellem DNA og transfektionsreagens er generelt anbefales at være 1:2-1:3.
(4) Sp.: Behøver det at blive afsluttet efter transfektion?
A: Ikke nødvendigt. Liposomkomplekser er stabile i 6 timer. Hvis cellemediet ikke ændres før transfektion, for at sikre de nødvendige næringsstoffer til normal cellevækst, er det nødvendigt at skifte til et nyt medium efter 4 til 6 timer. Men hvis mediet er blevet skiftet før transfektion, er det ikke nødvendigt at ændre mediet efter liposomtransfektion.
(5) Sp: Hvad skal jeg være opmærksom på, hvis jeg ønsker at forbedre transfektionseffektiviteten?
A: a: Tætheden af celler på tidspunktet for transfektion er 90%-95%.
b: Under transfektion, brug MEM-serumfrit medium til nukleinsyre- og liposomfortyndinger.
c: Mediet kan ændres 4-6 timer efter transfektion.
(6) Q: Kan co-transfektion af DNA og siRNA udføres? Hvordan er effekten?
A: Co-transfektion kan udføres, men det anbefales at udføre separat transfektion, og DNA-transfektion bør udføres 6 timer efter siRNA. Hvis det opereres sammen, vil siRNA-transfektionseffektiviteten være værre.
(7) Sp.: Kan transfektionsreagenset bruges til lentiviral emballagetransfektion?
A: Lentiviral emballage er mulig, men effektiviteten af lentiviral emballage er ikke nødvendigvis relateret til effektiviteten af transfektion, men også relateret til valget af emballageplasmider og forholdet mellem plasmider.
(8) Sp.: Kan Hieff Trans™ liposomnukleinsyretransfektionsreagens bruges til transfektion af suspensionsceller?
A: Hieff Trans™ liposomnukleinsyretransfektionsreagens kan bruges til suspensionscelletransfektion, se protokol for detaljer. Derudover har vi også lanceret et transfektionsreagens specifikt til suspensionsceller (Kat. nr. 40805, liposomnukleinsyretransfektionsreagens til suspensionsceller).
[1] Liu R, Yang J, Yao J, Zhao Z, He W, Su N, Zhang Z, Zhang C, Zhang Z, Cai H, Zhu L, Zhao Y, Quan S, Chen X, Yang Y. Optogenetisk kontrol af RNA-funktion og metabolisme ved hjælp af konstruerede lys-omskiftelige RNA-bindende proteiner. Nat Biotechnol. 2022 3. januar doi: 10.1038/s41587-021-01112-1. Epub forud for tryk. PMID: 34980910. (IF:54.908)
[2] Zhou J, Chen P, Wang H, Liu H, Li Y, Zhang Y, Wu Y, Paek C, Sun Z, Lei J, Yin L. Cas12a-varianter designet til lavere genomomfattende off-target-effekt gennem stringent PAM-genkendelse. Mol Ther. 2022 Jan 5;30(1):244-255. doi: 10.1016/j.ymthe.2021.10.010. Epub 2021 20. okt. PMID: 34687846; PMCID: PMC8753454. (IF:11.454)
[3] Chen S, Cao X, Zhang J, Wu W, Zhang B, Zhao F. circVAMP3 driver CAPRIN1-faseseparation og hæmmer hepatocellulært karcinom ved at undertrykke c-Myc-oversættelse. Adv Sci (Weinh).Marts 2022;9(8):e2103817. doi: 10.1002/advs.202103817. Epub 2022 Jan 24. PMID: 35072355; PMCID: PMC8922094. (IF:16.808)
[4] Zhang Y, Yu X, Sun R, Min J, Tang X, Lin Z, Xie S, Li X, Lu S, Tian Z, Gu C, Teng L, Yang Y. Splejsningsfaktor arginin/serin-rig 8 fremmer multipel myelom malignitet og knoglelæsion gennem alternativ splejsning af CACYBP og exosom-baseret cellulær kommunikation. Clin Transl Med. 2022 Feb;12(2):e684. doi: 10.1002/ctm2.684. PMID: 35184390. (IF:11.492)
[5] Tang X, Deng Z, Ding P, Qiang W, Lu Y, Gao S, Hu Y, Yang Y, Du J, Gu C. Et nyt protein kodet af circHNRNPU fremmer myelomatose progression ved at regulere knoglemarvens mikromiljø og alternativ splejsning. J Exp Clin Cancer Res. 2022 8. marts;41(1):85. doi: 10.1186/s13046-022-02276-7. PMID: 35260179. (IF:11.161)
[6] Hua Z, Wei R, Guo M, Lin Z, Yu X, Li X, Gu C, Yang Y. YTHDF2 fremmer multipelt myelomcelleproliferation via STAT5A/MAP2K2/p-ERK-aksen. Onkogen. 2022 Mar;41(10):1482-1491. doi: 10.1038/s41388-022-02191-3. Epub 2022 24. januar. PMID: 35075244. (IF:9.867)
[7] Liang Y, Lu Q, Li W, Zhang D, Zhang F, Zou Q, Chen L, Tong Y, Liu M, Wang S, Li W, Ren X, Xu P, Yang Z, Dong S, Zhang B, Huang Y, Li D, Wang H, Yu W. Reaktivering af tumorsuppressor i brystkræft ved forstærker-switch gennem NamiRNA-netværk. Nucleic Acids Res. 2021 7. september;49(15):8556-8572. doi: 10.1093/nar/gkab626. PMID: 34329471; PMCID: PMC8421228. (IF:16,9)
[8] Dai L, Dai Y, Han J, Huang Y, Wang L, Huang J, Zhou Z. Strukturel indsigt i BRCA1-BARD1 kompleks rekruttering til beskadiget kromatin. Mol Cell. 2021 Jul 1;81(13):2765-2777.e6. doi: 10.1016/j.molcel.2021.05.010. Epub 2021 7. juni. PMID: 34102105. (IF:17.97)
[9] Zhang K, Wang A, Zhong K, Qi S, Wei C, Shu X, Tu WY, Xu W, Xia C, Xiao Y, Chen A, Bai L, Zhang J, Luo B, Wang W, Shen C. UBQLN2-HSP70-aksen reducerer poly-Gly-Ala-modeller og lindrer adfærds-C9-defekter7ORF2. Neuron. 2021 Jun 16;109(12):1949-1962.e6. doi: 10.1016/j.neuron.2021.04.023. Epub 2021 14. maj. PMID: 33991504. (IF:17.17)
[10] Liang Y, Lu Q, Li W, Zhang D, Zhang F, Zou Q, Chen L, Tong Y, Liu M, Wang S, Li W, Ren X, Xu P, Yang Z, Dong S, Zhang B, Huang Y, Li D, Wang H, Yu W. Reaktivering af tumorsuppressor i brystkræft ved forstærker-netværksskifte gennem NamiRNA-netværk. Nucleic Acids Res. 2021 7. september;49(15):8556-8572. doi: 10.1093/nar/gkab626. PMID: 34329471; PMCID: PMC8421228. (IF:16,9)
[11] Li T, Chen X, Qian Y, Shao J, Li X, Liu S, Zhu L, Zhao Y, Ye H, Yang Y. En syntetisk BRET-baseret optogenetisk enhed til pulserende transgenekspression, der muliggør glukosehomeostase i mus. Nat Commun. 2021 Jan 27;12(1):615. doi: 10.1038/s41467-021-20913-1. PMID: 33504786; PMCID: PMC7840992. (IF:14,92)
[12] Pan Y, He X, Li C, Li Y, Li W, Zhang H, Wang Y, Zhou G, Yang J, Li J, Qu J, Wang H, Gao Z, Shen Y, Li T, Hu H, Ma H. Neuronal aktivitet rekrutterer CRTC1/CREB-aksen til at drive transskriptionsafhængig autofagi LTD. Cell Rep. 2021 Jul 20;36(3):109398. doi: 10.1016/j.celrep.2021.109398. PMID: 34289350. (IF:9.42)
[13] Liu H, Xing R, Ou Z, Zhao J, Hong G, Zhao TJ, Han Y, Chen Y. G-proteinkoblet receptor GPR17 hæmmer gliomudvikling ved at øge polycomb-repressivt kompleks 1-medieret ROS-produktion. Celledød Dis. 12. juni 2021;12(6):610. doi: 10.1038/s41419-021-03897-0. PMID: 34120140; PMCID: PMC8197764. (IF:8.463)
[14] Fan Y, Wang J, Jin W, Sun Y, Xu Y, Wang Y, Liang X, Su D. CircNR3C2 fremmer HRD1-medieret tumor-undertrykkende effekt via sponging miR-513a-3p i triple-negativ brystkræft. Mol Cancer. 2. februar 2021; 20(1):25. doi: 10.1186/s12943-021-01321-x.PMID: 33530981; PMCID: PMC7851937. (IF:27.403)
[15] Gu C, Wang Y, Zhang L, Qiao L, Sun S, Shao M, Tang X, Ding P, Tang C, Cao Y, Zhou Y, Guo M, Wei R, Li N, Xiao Y, Duan J, Yang Y. AHSA1 er et lovende terapeutisk mål for cellulær multipel proliferation og proteasomhæmmerresistens. J Exp Clin Cancer Res. 2022 6. januar;41(1):11. doi: 10.1186/s13046-021-02220-1. PMID: 34991674; PMCID: PMC8734095. (IF:11.161)
[16] Luo Q, Wu X, Zhao P, Nan Y, Chang W, Zhu X, Su D, Liu Z. OTUD1 Aktiverer Caspase-uafhængig og Caspase-afhængig apoptose ved at fremme AIF-kernetranslokation og MCL1-nedbrydning. Adv Sci (Weinh). 8. februar 2021; 8(8):2002874. doi: 10.1002/advs.202002874. PMID: 33898171; PMCID: PMC8061361. (IF:15,84)
[17] Luo Q, Wu X, Nan Y, Chang W, Zhao P, Zhang Y, Su D, Liu Z. TRIM32/USP11 balancerer ARID1A-stabilitet og den onkogene/tumor-suppressive status af pladecellekarcinom. Cell Rep. 2020 Jan 7;30(1):98-111.e5. doi: 10.1016/j.celrep.2019.12.017. PMID: 31914402. (IF:9.42)
[18] Sun X, Peng X, Cao Y, Zhou Y, Sun Y. ADNP fremmer neural differentiering ved at modulere Wnt/β-catenin signalering. Nat Commun. 12. juni 2020;11(1):2984. doi: 10.1038/s41467-020-16799-0. PMID: 32533114; PMCID: PMC7293280. (IF:14.911)
[19] Yang X, Wang H, Xie E, Tang B, Mu Q, Song Z, Chen J, Wang F, Min J. Omledning af ERBB3- og ERK-signalering bibringer resistens mod FGFR1-hæmning i mave-tarmkræft med en ERBB3-E928G-mutation. Proteincelle. 2020 Dec;11(12):915-920. doi: 10.1007/s13238-020-00749-z. PMID: 32632529; PMCID: PMC7719122. (IF:14.872)
[20] Chen, T., Chen, Y., Chen, H. et al. Dual-enzym-drevet ubundet DNA walking nanomaskine til intracellulær billeddannelse af lavt udtrykt mikroRNA. Nano Res. 12, 1055-1060 (2019). https://doi.org/10.1007/s12274-019-2344-5 (IF:8.21)
[21] Zhang X, Qi Z, Yin H, Yang G. Interaktion mellem p53 og Ras-signalering kontrollerer cisplatinresistens via HDAC4- og HIF-1α-medieret regulering af apoptose og autofagi. Teranostik. 2019 Jan 30;9(4):1096-1114. doi: 10.7150/thno.29673. PMID: 30867818; PMCID: PMC6401400. (IF:8.12)
[22] Zou Y, Wang A, Shi M, Chen X, Liu R, Li T, Zhang C, Zhang Z, Zhu L, Ju Z, Loscalzo J, Yang Y, Zhao Y. Analyse af redoxlandskaber og dynamik i levende celler og in vivo ved hjælp af genetisk kodede fluorescerende sensorer. Nat Protoc. 2018 okt;13(10):2362-2386. doi: 10.1038/s41596-018-0042-5. PMID: 30258175; PMCID: PMC6714056. (IF:13,49)
[23] Zhang K, Zhao X, Chen X, Wei Y, Du W, Wang Y, Liu L, Zhao W, Han Z, Kong D, Zhao Q, Guo Z, Han Z, Liu N, Ma F, Li Z. Forstærkede terapeutiske virkninger af mesenkymale stamcelle-afledte exosomer med en injicerbar hydrokemi til injicerbar hydrogel. ACS Appl Mater-grænseflader. 12. september 2018;10(36):30081-30091. doi: 10.1021/acsami.8b08449. Epub 2018 29. august. PMID: 30118197. (IF:8.09)
[24] Hao H, Hu S, Chen H, Bu D, Zhu L, Xu C, Chu F, Huo X, Tang Y, Sun X, Ding BS, Liu DP, Hu S, Wang M. Tab af endotel CXCR7 forringer vaskulær homeostase og hjerteombygning efter myokardieinfarkt Drug for hjerteinfarkt: implikationer. Cirkulation. 2017 Mar 28;135(13):1253-1264. doi: 10.1161/CIRCULATIONAHA.116.023027. Epub 2017 2. februar PMID: 28154007. (IF:18.881)
Betaling og sikkerhed
Dine betalingsoplysninger behandles sikkert. Vi gemmer ikke kreditkortoplysninger og har heller ikke adgang til dine kreditkortoplysninger.
Forespørgsel
Du kan også lide
FAQ
Produktet er kun til forskningsformål og er ikke beregnet til terapeutisk eller diagnostisk brug hos mennesker eller dyr. Produkter og indhold er beskyttet af patenter, varemærker og ophavsrettigheder ejet af Yeasen Biotechnology. Varemærkesymboler angiver oprindelseslandet, ikke nødvendigvis registrering i alle regioner.
Visse applikationer kan kræve yderligere tredjeparts intellektuelle ejendomsrettigheder.
Yeasen er dedikeret til etisk videnskab og mener, at vores forskning bør adressere kritiske spørgsmål og samtidig sikre sikkerhed og etiske standarder.