


Descripción
La mezcla maestra para PCR 2×Hieff™ contiene la polimerasa de ADN Taq Hieff™ (Cat. n.° 10101), dNTP y otros componentes necesarios para la PCR. La mezcla maestra es estable durante 3 meses a 4 °C con nuestros estabilizadores personalizados. La solución de premezcla está optimizada para la PCR convencional y está lista para usar al agregar la plantilla de ADN y los cebadores. Los productos de PCR se pueden cargar directamente para la electroforesis con el colorante azul de bromofenol precargado. Los productos amplificados contienen una protrusión de 3'-dA y se pueden clonar fácilmente en el vector T. La mezcla maestra para PCR 2×Hieff™ simplifica el procedimiento de PCR y reduce la contaminación.
Características
- Mezcla conveniente y lista para usar
- Termoestable: la vida media es de más de 40 minutos a 95 °C.
- Genera productos de PCR con salientes 3'-dA
Aplicaciones
- Amplificación de rutina de fragmentos de ADN mediante PCR
- PCR de alto rendimiento
- Etiquetado de ADN
Presupuesto
| Fidelidad (contra Taq) | 1 × |
| Arranque en caliente | No |
| Sobresalir | 3'-A |
| Polimerasa | Taq ADN polimerasa |
| Formato de reacción | SuperMix o mezcla maestra |
| Velocidad de reacción | Estándar |
| Tipo de producto | Mezcla maestra de PCR (2×) |
Componentes
| Componentes No. | Nombre | 10102ES03 | 10102ES08 | 10102ES50 | 10102ES60 |
| 10102 | 2×Hief™ Mezcla maestra de PCR (con colorante) | 1 ml | 5 x 1 ml | 50 x 1 ml | 100×1 ml |
Envío y almacenamiento
El 2×Hieff™ Los productos PCR Master Mix deben almacenarse a -15 ℃ ~-25℃ por 2 años.
Cifras
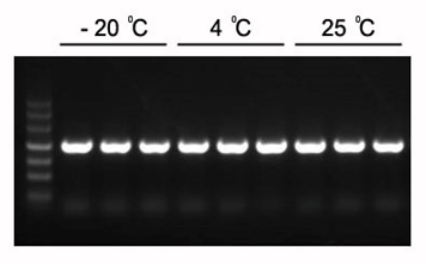
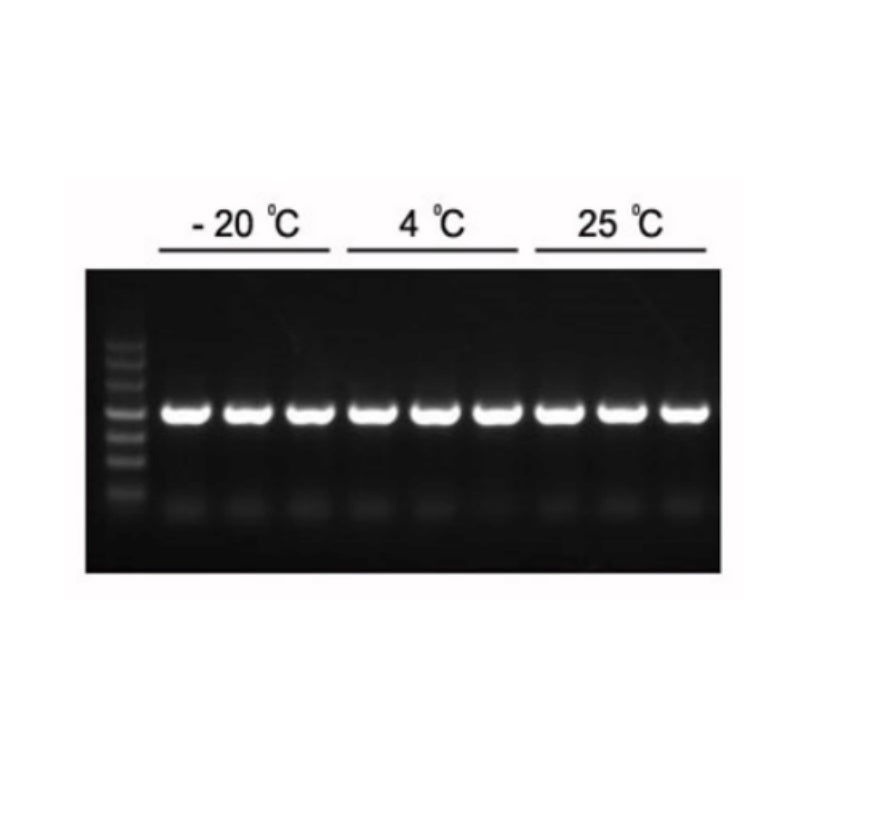
Figura 1. Los productos de PCR esperados de 1,2 kb se pueden amplificar con 2×Hieff PCR Master Mix.
La mezcla maestra se almacenó a -20 °C durante 1 año, después se almacenó durante 3 meses a 4 °C y 1 mes a 25 °C. Plantilla: genoma de Arabidopsis. Temperatura de hibridación: 60 °C. Tiempo de extensión: 40 s.
Citado de "Sensores ultrasensibles revelan el panorama espaciotemporal del metabolismo del lactato en fisiología y enfermedad"
Cell Metab. 3 de enero de 2023;35(1):200-211.e9. doi: 10.1016/j.cmet.2022.10.002
[1] Wu LY, Shang GD, Wang FX, et al. El perfil dinámico del estado de la cromatina revela funciones reguladoras de la auxina y la citoquinina en la regeneración de los brotes. Dev Cell. 2022;57(4):526-542.e7. doi:10.1016/j.devcel.2021.12.019(IF:12.270)
[2] Li Y, Wang C, Zhang L, Chen B, Mo Y, Zhang J. La claudin-5a es esencial para la formación funcional de la barrera hematoencefálica y la barrera hematoencefálica del pez cebra. Fluids Barriers CNS. 2022;19(1):40. Publicado el 3 de junio de 2022. doi:10.1186/s12987-022-00337-9(IF:7.662)
[3] Luo T, Wang Y, Tang H, et al. Una terapia génica dirigida a NF-κB basada en AAV (rAAV-DMP-miR533) para enfermedades inflamatorias. J Inflamm Res. 2022;15:3447-3466. Publicado el 14 de junio de 2022. doi:10.2147/JIR.S362732(IF:6.922)
[4] Chen Z, Qi Z, He D, et al. Estrategia para escanear ARN circulares codificadores de péptidos en cáncer colorrectal basada en análisis bioinformáticos y ensayos experimentales. Front Cell Dev Biol. 2022;9:815895. Publicado el 25 de febrero de 2022. doi:10.3389/fcell.2021.815895(IF:6.684)
[5] Yu Y, Fang L. CircRPAP2 regula el empalme alternativo de PTK2 al unirse a SRSF1 en el cáncer de mama. Cell Death Discov. 2022;8(1):152. Publicado el 2 de abril de 2022. doi:10.1038/s41420-022-00965-y(IF:5.241)
[6] Zhang J, Liu W, Li G, et al. BCAS2 está involucrado en el splicing alternativo y el desarrollo de ovocitos de ratón. FASEB J. 2022;36(2):e22128. doi:10.1096/fj.202101279R(IF:5.192)
[7] Xiao W, Li J, Zhang Y, et al. Una cepa fúngica de Bipolaris bicolor como posible bioherbicida para el control de la hierba de ganso (Eleusine indica). Pest Manag Sci. 2022;78(3):1251-1264. doi:10.1002/ps.6742(IF:4.845)
[8] Zhang Y, Yu R, Tang J, et al. Tres genes de la familia CYP4 del citocromo P450 regulados por la vía de señalización CncC median la susceptibilidad fitoquímica en el gorgojo rojo de la harina, Tribolium castaneum. Pest Manag Sci. 2022;78(8):3508-3518. doi:10.1002/ps.6991(IF:4.845)
[9] Zhang X, Yang S, Chen W, et al. Circular RNA circYPEL2: un nuevo biomarcador en el cáncer de cuello uterino. Genes (Basilea). 2021;13(1):38. Publicado el 23 de diciembre de 2021. doi:10.3390/genes13010038(IF:4.096)
[10] Gu K, Qian D, Qin H, et al. Una nueva mutación en KCNH2 produce pérdida de función del canal de potasio hERG en el síndrome de QT largo 2. Pflugers Arch. 2021;473(2):219-229. doi:10.1007/s00424-021-02518-1(IF:3.657)
[11] Yang Y, Chu X, Nie M, et al. Una nueva deleción de largo alcance que abarca STX16 y NPEPL1 que causa defectos de impronta del locus GNAS descubiertos en un paciente con pseudohipoparatiroidismo autosómico dominante tipo 1B. Endocrine. 2020;69(1):212-219. doi:10.1007/s12020-020-02304-6(IF:3.235)
[12] Ullah H, Arbab S, Khan MIU, et al. Fragmento de ADN mitocondrial circulante libre de células: un posible marcador para la detección temprana de Schistosoma japonicum. Infect Genet Evol. 2021;88:104683. doi:10.1016/j.meegid.2020.104683(IF:2.773)
[13] Xu L, Chen Y, Shen T, Lin C, Zhang B. Análisis genético del gen PICK1 en la enfermedad de Alzheimer: un estudio para encontrar un nuevo gen diana. Front Neurol. 2019;9:1169. Publicado el 9 de enero de 2019. doi:10.3389/fneur.2018.01169(IF:2.635)
[14] Ullah H, Qadeer A, Giri BR. Detección de ADN circulante libre de células para diagnosticar la infección por Schistosoma japonicum. Acta Trop. 2020;211:105604. doi:10.1016/j.actatropica.2020.105604(IF:2.555)
[15] Yan ZC, Hua HQ, Qi GY, Li YX. Detección temprana e identificación de avispas parasitoides Trichogramma Westwood (Hymenoptera: Trichogrammatidae) en sus huevos hospedadores mediante polimorfismo de longitud de fragmentos de restricción de reacción en cadena de la polimerasa [publicado en línea antes de su impresión, 24 de junio de 2022]. J Econ Entomol. 2022;toac095. doi:10.1093/jee/toac095(IF:2.381)
[16] Zhou Z, Yin H, Suye S, Zhu F, Cai H, Fu C. Cambios en las roturas de doble cadena de ADN y la reparación del ADN durante la formación de la reserva ovárica en ratones. Reprod Biol. 2022;22(1):100603. doi:10.1016/j.repbio.2022.100603(IF:2.376)
Pago y seguridad
Su información de pago se procesa de forma segura. No almacenamos detalles de la tarjeta de crédito ni tenemos acceso a la información de su tarjeta de crédito.
Consulta
También te puede gustar
Preguntas frecuentes
El producto es solo para fines de investigación y no está destinado a uso terapéutico o diagnóstico en humanos o animales. Los productos y el contenido están protegidos por patentes, marcas comerciales y derechos de autor propiedad de Yeasen Biotechnology. Los símbolos de marca comercial indican el país de origen, no necesariamente el registro en todas las regiones.
Algunas aplicaciones pueden requerir derechos de propiedad intelectual adicionales de terceros.
Yeasen se dedica a la ciencia ética y cree que nuestra investigación debe abordar cuestiones críticas al tiempo que garantiza la seguridad y los estándares éticos.