


Descripción
Hieff Trans™ El Reactivo de Transfección Liposomal es un reactivo versátil para la transfección de liposomas, apto para la transfección de ADN, ARN y oligonucleótidos, con una alta eficiencia de transfección para la mayoría de las células eucariotas. Su fórmula única permite añadirlo directamente al medio, y la presencia de suero no afecta la eficiencia de la transfección, lo que reduce el daño celular causado por la eliminación del suero. No es necesario eliminar el complejo de ácido nucleico-Hieff Trans™ ni reemplazarlo con un medio nuevo después de la transfección, y también puede eliminarse después de 4-6 horas.
Hieff Trans™ Se suministra en forma líquida estéril. Normalmente, para la transfección en placas de 24 pocillos, se utilizan aproximadamente 1,5 μL cada vez, 1 mL de Hieff Trans™. puede realizar aproximadamente 660 transfecciones; para una placa de 6 pocillos, aproximadamente 6 μL cada vez, 1 mL de Hieff Trans™ puede realizar aproximadamente 660 transfecciones. 160 transfecciones.
Característica
- Eficiencia excepcional: Rendimiento de transfección superior en una amplia gama de líneas celulares, incluida la transfección transitoria y la transfección estable.
- Amplia adaptabilidad: Excelente eficiencia de transfección en una variedad de líneas celulares y altos niveles de expresión de proteína recombinante.
- Baja toxicidad: Las actividades de las diversas células casi no se ven afectadas por los reactivos de transfección de
YEASEN - Operación simple: eficacia comprobada en presencia de suero, lo que elimina la necesidad de cambiar el medio después de la transfección.
- Rentable: Efecto de transfección competitivo con precios más asequibles
Solicitud
- Transfección celular
Especificación
| Forma | Líquido |
| Compatible con suero | Sí |
| Tipo de célula | Líneas celulares establecidas |
| Tipo de muestra | ADN plasmídico, ARNi sintético |
| Técnica de transfección | Transfección basada en lípidos |
Componentes
| Componentes No. | Nombre | 40802ES02 | 40802ES03 | 40802ES08 |
| 40802 | Hieff Trans™ Reactivo de transfección liposomal | 0,5 ml | 1 ml | 5×1 ml |
Envío y almacenamiento
El producto se envía con bolsas de hielo y puede conservarse a una temperatura de 2 a 8 °C durante un año. ¡No congelar!
Cifras

Citado de: Biología de las Comunicaciones, volumen 5, número de artículo: 1296 (2022)
- Demostración del efecto de transfección
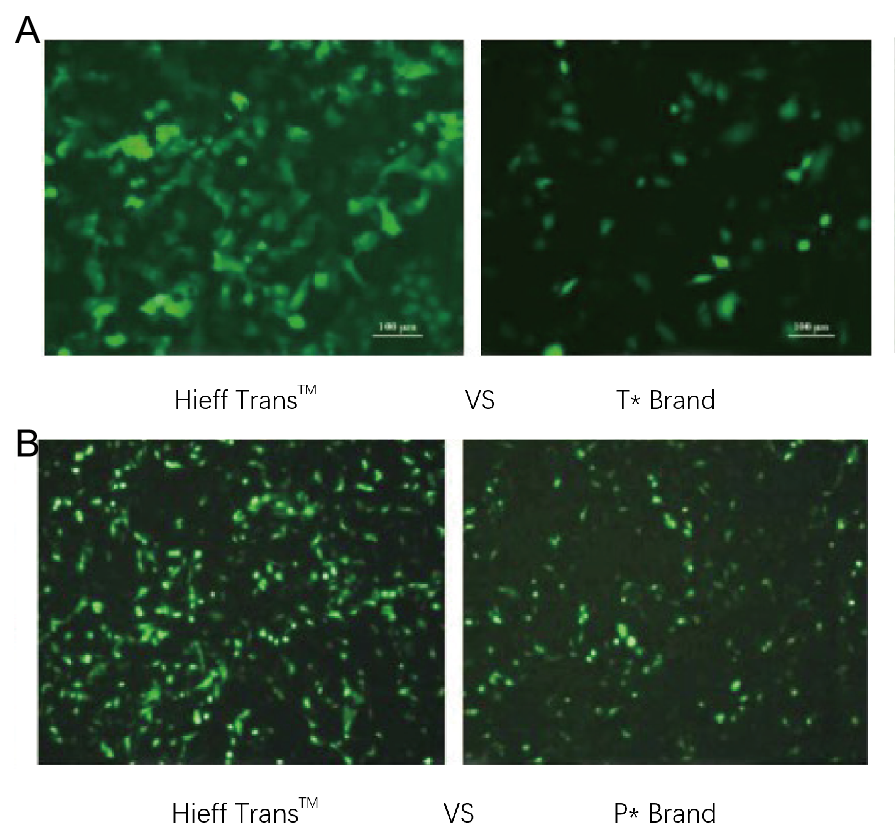
Figura 1. El reactivo de transfección de liposomas Hieff Trans™ supera al reactivo de transfección de la marca competitiva (marca T* en la Figura 1a, marca P* en la Figura 1b).
Cada reactivo se utilizó para transfectar la línea celular diana (línea celular Hela en la Figura 1a, línea celular DF-1 en la Figura 1b) en un formato de 96 pocillos. La expresión de GFP se analizó 48 horas después de la transfección. El reactivo de transfección de liposomas Hieff Trans™ proporcionó una mayor eficiencia de transfección de GFP que los productos de la competencia.
- Líneas celulares validadas
| Producto Nombre | Hieff TransTMLiposomal Transfección Reactivo | |||||
| Validado Celúla Pauta | 293T | Calu 1 | HEK293 | HO1980 | N2A | TS |
| 293F | CHO-K1 | HEK293T | HUVEC | NCI-H1975 | U-87 | |
| 293 pies | COS-7 | Hola | MCF10A | NIH-3T3 | Vero | |
| 3T3 | DF-1 | Hep2C | MCF-7 | Neuro-2a | WEHI | |
| A549 | H1299 | Hep3B | MDA-MB-231 | PC-12 | WRL-68 | |
| BV-2 | H520 | Hepa1-6 | MDA-MB-231-LM2-4175 | Real academia de bellas artesw264.7 | ||
| B50 | HaCaT | HepG2 | MDCK | SGC-7901 | ||
| C2C12 | HCT116 | HK2 | Ministerio de Economía y Finanzas | T47D | … | |
Figura 2. Lista de líneas celulares transfectadas con éxito con Hieff Trans™ Reactivo de Transfección Liposomal (En continua actualización).

Citado de "Control optogenético de la función y el metabolismo del ARN mediante proteínas de unión al ARN conmutables por luz diseñadas". Nat Biotechnol. 3 de enero de 2022. doi: 10.1038/s41587-021-01112-1"

Citado de "Información estructural sobre el reclutamiento del complejo BRCA1-BARD1 a la cromatina dañada". Mol Cell. 1 de julio de 2021;81(13):2765-2777.e6. doi: 10.1016/j.molcel.2021.05.010.
Citado de "El eje UBQLN2-HSP70 reduce los agregados de poli-Gly-Ala y alivia los defectos de comportamiento en el modelo animal C9ORF72". Neuron. 16 de junio de 2021;109(12):1949-1962.e6. doi: 10.1016/j.neuron.2021.04.023.
Hieff Trans™ Reactivo de transfección liposomal Preguntas frecuentes
(1) P: ¿Puede haber suero presente al preparar el complejo de reactivo de transfección de ácidos nucleicos?
R: La presencia de suero afecta la formación de liposomas. Se recomienda utilizar un medio sin suero (generalmente medio MEM) al preparar complejos de reactivos de transfección de ácidos nucleicos.
(2) P: ¿Se puede congelar el reactivo de transfección?
R: No. Este reactivo debe almacenarse a una temperatura de 2 a 8 °C y se debe tener cuidado de no abrir la tapa repetidamente durante un tiempo prolongado, ya que la apertura prolongada de la tapa provocará la oxidación de los liposomas y afectará la eficiencia de la transfección.
(3) P: ¿A qué debo prestar atención al utilizar el reactivo de transfección de ácidos nucleicos liposomales Hieff Trans™?
R: 1) Durante la operación de transfección, es mejor que la confluencia celular alcance el 80%-95%, y la densidad de siembra específica se determina de acuerdo con la situación de las células;
2) El uso de ADN de alta pureza ayuda a obtener una mayor eficiencia de transfección;
3) Es necesario diluir el ADN y los reactivos de transfección con medio sin suero cuando se preparan complejos de transfección;
4) No se pueden añadir antibióticos al medio durante la transfección;
5) La concentración de ADN y la cantidad de reactivo liposomal catiónico deben optimizarse para el primer uso a fin de obtener la máxima eficiencia de transfección. La proporción de ADN a reactivo de transfección es Generalmente se recomienda que sea 1:2-1:3.
(4) P: ¿Es necesario finalizarlo después de la transfección?
R: No es necesario. Los complejos liposomales son estables durante 6 horas. Si no se modifica el medio celular antes de la transfección, para asegurar los nutrientes necesarios para el crecimiento celular normal, es necesario cambiarlo después de 4 a 6 horas. Sin embargo, si se modificó el medio antes de la transfección, no es necesario cambiarlo después de la transfección de liposomas.
(5) P: ¿A qué debo prestar atención si quiero mejorar la eficiencia de la transfección?
R: a: La densidad de células en el momento de la transfección es del 90%-95%.
b: Durante la transfección, utilice medio MEM sin suero para diluciones de ácidos nucleicos y liposomas.
c: El medio se puede cambiar 4-6 horas después de la transfección.
(6) P: ¿Se puede realizar la cotransfección de ADN y ARNi? ¿Cuál es el efecto?
R: Se puede realizar la cotransfección, pero se recomienda realizarla por separado. La transfección de ADN debe realizarse 6 horas después de la del ARNip. Si se realizan conjuntamente, la eficiencia de la transfección del ARNip será menor.
(7) P: ¿Se puede utilizar el reactivo de transfección para la transfección de paquetes lentivirales?
R: El empaquetamiento lentiviral es posible, pero la eficiencia del empaquetamiento lentiviral no está necesariamente relacionada con la eficiencia de la transfección, sino también con la selección de plásmidos de empaquetamiento y la proporción entre plásmidos.
(8) P: ¿Se puede utilizar el reactivo de transfección de ácidos nucleicos liposomales Hieff Trans™ para la transfección de células en suspensión?
R: El reactivo de transfección de ácidos nucleicos liposomales Hieff Trans™ puede utilizarse para la transfección de células en suspensión. Consulte el protocolo para obtener más información. Además, hemos lanzado un reactivo de transfección específico para células en suspensión (n.° de cat. 40805, reactivo de transfección de ácidos nucleicos liposomales para células en suspensión).
[1] Liu R, Yang J, Yao J, Zhao Z, He W, Su N, Zhang Z, Zhang C, Zhang Z, Cai H, Zhu L, Zhao Y, Quan S, Chen X, Yang Y. Control optogenético de la función y el metabolismo del ARN mediante proteínas de unión al ARN conmutables por luz, diseñadas. Nat Biotechnol. 3 de enero de 2022. doi: 10.1038/s41587-021-01112-1. Publicación electrónica previa a la impresión. PMID: 34980910. (IF:54.908)
[2] Zhou J, Chen P, Wang H, Liu H, Li Y, Zhang Y, Wu Y, Paek C, Sun Z, Lei J, Yin L. Variantes de Cas12a diseñadas para reducir el efecto fuera del objetivo en todo el genoma mediante el reconocimiento riguroso de PAM. Mol Ther. 5 de enero de 2022;30(1):244-255. doi: 10.1016/j.ymthe.2021.10.010. Publicación electrónica: 20 de octubre de 2021. PMID: 34687846; PMCID: PMC8753454. (IF:11.454)
[3] Chen S, Cao X, Zhang J, Wu W, Zhang B, Zhao F. circVAMP3 impulsa la separación de fases de CAPRIN1 e inhibe el carcinoma hepatocelular al suprimir la traducción de c-Myc. Adv Sci (Weinh).Marzo de 2022;9(8):e2103817. doi: 10.1002/advs.202103817. Publicación electrónica, 24 de enero de 2022. PMID: 35072355; PMCID: PMC8922094. (IF:16.808)
[4] Zhang Y, Yu X, Sun R, Min J, Tang X, Lin Z, Xie S, Li X, Lu S, Tian Z, Gu C, Teng L, Yang Y. El factor de empalme 8 rico en arginina/serina promueve la malignidad del mieloma múltiple y la lesión ósea mediante el empalme alternativo de CACYBP y la comunicación celular basada en exosomas. Clin Transl Med. Febrero de 2022;12(2):e684. doi: 10.1002/ctm2.684. PMID: 35184390. (IF:11.492)
[5] Tang X, Deng Z, Ding P, Qiang W, Lu Y, Gao S, Hu Y, Yang Y, Du J, Gu C. Una nueva proteína codificada por circHNRNPU promueve la progresión del mieloma múltiple mediante la regulación del microambiente de la médula ósea y el empalme alternativo. J Exp Clin Cancer Res. 8 de marzo de 2022; 41 (1): 85. doi: 10.1186/s13046-022-02276-7. PMID: 35260179. (SI:11.161)
[6] Hua Z, Wei R, Guo M, Lin Z, Yu X, Li X, Gu C, Yang Y. YTHDF2 promueve la proliferación de células de mieloma múltiple mediante el eje STAT5A/MAP2K2/p-ERK. Oncogene. Marzo de 2022;41(10):1482-1491. doi: 10.1038/s41388-022-02191-3. Publicación electrónica, 24 de enero de 2022. PMID: 35075244. (IF:9.867)
[7] Liang Y, Lu Q, Li W, Zhang D, Zhang F, Zou Q, Chen L, Tong Y, Liu M, Wang S, Li W, Ren X, Xu P, Yang Z, Dong S, Zhang B, Huang Y, Li D, Wang H, Yu W. Reactivación del supresor tumoral en cáncer de mama mediante conmutación de potenciador a través de la red NamiRNA. Nucleic Acids Res. 7 de septiembre de 2021;49(15):8556-8572. doi: 10.1093/nar/gkab626. PMID: 34329471; PMCID: PMC8421228. (SI:16.9)
[8] Dai L, Dai Y, Han J, Huang Y, Wang L, Huang J, Zhou Z. Perspectiva estructural del reclutamiento del complejo BRCA1-BARD1 a la cromatina dañada. Mol Cell. 1 de julio de 2021;81(13):2765-2777.e6. doi: 10.1016/j.molcel.2021.05.010. Publicación electrónica: 7 de junio de 2021. PMID: 34102105. (IF:17.97)
[9] Zhang K, Wang A, Zhong K, Qi S, Wei C, Shu X, Tu WY, Xu W, Xia C, Xiao Y, Chen A, Bai L, Zhang J, Luo B, Wang W, Shen C. El eje UBQLN2-HSP70 reduce los agregados de poli-Gly-Ala y alivia los defectos de comportamiento en el modelo animal C9ORF72. Neuron. 16 de junio de 2021;109(12):1949-1962.e6. doi: 10.1016/j.neuron.2021.04.023. Publicación electrónica: 14 de mayo de 2021. PMID: 33991504. (IF:17.17)
[10] Liang Y, Lu Q, Li W, Zhang D, Zhang F, Zou Q, Chen L, Tong Y, Liu M, Wang S, Li W, Ren X, Xu P, Yang Z, Dong S, Zhang B, Huang Y, Li D, Wang H, Yu W. Reactivación del supresor tumoral en cáncer de mama mediante la conmutación de potenciadores a través de la red NamiRNA. Nucleic Acids Res. 7 de septiembre de 2021;49(15):8556-8572. doi: 10.1093/nar/gkab626. PMID: 34329471; PMCID: PMC8421228. (IF:16.9)
[11] Li T, Chen X, Qian Y, Shao J, Li X, Liu S, Zhu L, Zhao Y, Ye H, Yang Y. Un dispositivo optogenético sintético basado en BRET para la expresión pulsátil de transgenes que permite la homeostasis de la glucosa en ratones. Nat Commun. 27 de enero de 2021;12(1):615. doi: 10.1038/s41467-021-20913-1. PMID: 33504786; PMCID: PMC7840992. (IF:14.92)
[12] Pan Y, He X, Li C, Li Y, Li W, Zhang H, Wang Y, Zhou G, Yang J, Li J, Qu J, Wang H, Gao Z, Shen Y, Li T, Hu H, Ma H. La actividad neuronal recluta el eje CRTC1/CREB para impulsar la autofagia dependiente de la transcripción y mantener la LTD en fase tardía. Cell Rep. 20 de julio de 2021;36(3):109398. doi: 10.1016/j.celrep.2021.109398. PMID: 34289350. (IF:9.42)
[13] Liu H, Xing R, Ou Z, Zhao J, Hong G, Zhao TJ, Han Y, Chen Y. El receptor acoplado a proteína G, GPR17, inhibe el desarrollo de gliomas al aumentar la producción de ROS mediada por el complejo represor Polycomb 1. Cell Death Dis. 12 de junio de 2021;12(6):610. doi: 10.1038/s41419-021-03897-0. PMID: 34120140; PMCID: PMC8197764. (IF:8.463)
[14] Fan Y, Wang J, Jin W, Sun Y, Xu Y, Wang Y, Liang X, Su D. CircNR3C2 promueve el efecto supresor tumoral mediado por HRD1 mediante la absorción de miR-513a-3p en el cáncer de mama triple negativo. Mol Cancer. 2 de febrero de 2021;20(1):25. doi: 10.1186/s12943-021-01321-x.PMID: 33530981; PMCID: PMC7851937. (IF:27.403)
[15] Gu C, Wang Y, Zhang L, Qiao L, Sun S, Shao M, Tang X, Ding P, Tang C, Cao Y, Zhou Y, Guo M, Wei R, Li N, Xiao Y, Duan J, Yang Y. AHSA1 es una diana terapéutica prometedora para la proliferación celular y la resistencia a los inhibidores del proteasoma en el mieloma múltiple. J Exp Clin Cancer Res. 6 de enero de 2022;41(1):11. doi: 10.1186/s13046-021-02220-1. PMID: 34991674; PMCID: PMC8734095. (IF:11.161)
[16] Luo Q, Wu X, Zhao P, Nan Y, Chang W, Zhu X, Su D, Liu Z. OTUD1 activa la apoptosis independiente y dependiente de caspasa al promover la translocación nuclear de AIF y la degradación de MCL1. Adv Sci (Weinh). 8 de febrero de 2021;8(8):2002874. doi: 10.1002/advs.202002874. PMID: 33898171; PMCID: PMC8061361. (IF:15.84)
[17] Luo Q, Wu X, Nan Y, Chang W, Zhao P, Zhang Y, Su D, Liu Z. TRIM32/USP11 equilibra la estabilidad de ARID1A y el estado oncogénico/supresor de tumores del carcinoma de células escamosas. Cell Rep. 7 de enero de 2020;30(1):98-111.e5. doi: 10.1016/j.celrep.2019.12.017. PMID: 31914402. (IF:9.42)
[18] Sun X, Peng X, Cao Y, Zhou Y, Sun Y. ADNP promueve la diferenciación neuronal mediante la modulación de la señalización de Wnt/β-catenina. Comuna Nacional. 12 de junio de 2020;11(1):2984. doi: 10.1038/s41467-020-16799-0. PMID: 32533114; PMCID: PMC7293280. (SI:14.911)
[19] Yang X, Wang H, Xie E, Tang B, Mu Q, Song Z, Chen J, Wang F, Min J. La reconexión de la señalización de ERBB3 y ERK confiere resistencia a la inhibición de FGFR1 en el cáncer gastrointestinal con mutación ERBB3-E928G. Protein Cell. Diciembre de 2020;11(12):915-920. doi: 10.1007/s13238-020-00749-z. PMID: 32632529; PMCID: PMC7719122. (IF:14.872)
[20] Chen, T., Chen, Y., Chen, H. et al. Nanomáquina de ADN móvil sin límite propulsada por enzimas duales para la obtención de imágenes intracelulares de microARN de baja expresión. Nano Res. 12, 1055–1060 (2019). https://doi.org/10.1007/s12274-019-2344-5 (SI:8.21)
[21] Zhang X, Qi Z, Yin H, Yang G. La interacción entre la señalización de p53 y Ras controla la resistencia al cisplatino mediante la regulación de la apoptosis y la autofagia mediada por HDAC4 y HIF-1α. Theranostics. 30 de enero de 2019;9(4):1096-1114. doi: 10.7150/thno.29673. PMID: 30867818; PMCID: PMC6401400. (IF:8.12)
[22] Zou Y, Wang A, Shi M, Chen X, Liu R, Li T, Zhang C, Zhang Z, Zhu L, Ju Z, Loscalzo J, Yang Y, Zhao Y. Análisis de paisajes redox y dinámica en células vivas e in vivo mediante sensores fluorescentes codificados genéticamente. Nat Protoc. Oct. 2018;13(10):2362-2386. doi: 10.1038/s41596-018-0042-5. PMID: 30258175; PMCID: PMC6714056. (IF:13.49)
[23] Zhang K, Zhao X, Chen X, Wei Y, Du W, Wang Y, Liu L, Zhao W, Han Z, Kong D, Zhao Q, Guo Z, Han Z, Liu N, Ma F, Li Z. Efectos terapéuticos mejorados de exosomas derivados de células madre mesenquimales con un hidrogel inyectable para el tratamiento de la isquemia de las extremidades posteriores. ACS Appl Mater Interfaces. 12 de septiembre de 2018;10(36):30081-30091. doi: 10.1021/acsami.8b08449. Publicación electrónica, 29 de agosto de 2018. PMID: 30118197. (IF:8.09)
[24] Hao H, Hu S, Chen H, Bu D, Zhu L, Xu C, Chu F, Huo X, Tang Y, Sun X, Ding BS, Liu DP, Hu S, Wang M. La pérdida de CXCR7 endotelial altera la homeostasis vascular y la remodelación cardíaca tras un infarto de miocardio: Implicaciones para el descubrimiento de fármacos cardiovasculares. Circulation. 28 de marzo de 2017;135(13):1253-1264. doi: 10.1161/CIRCULATIONAHA.116.023027. Publicación electrónica: 2 de febrero de 2017. PMID: 28154007. (IF:18.881)
Pago y seguridad
Su información de pago se procesa de forma segura. No almacenamos detalles de la tarjeta de crédito ni tenemos acceso a la información de su tarjeta de crédito.
Consulta
También te puede gustar
Preguntas frecuentes
El producto es solo para fines de investigación y no está destinado a uso terapéutico o diagnóstico en humanos o animales. Los productos y el contenido están protegidos por patentes, marcas comerciales y derechos de autor propiedad de Yeasen Biotechnology. Los símbolos de marca comercial indican el país de origen, no necesariamente el registro en todas las regiones.
Algunas aplicaciones pueden requerir derechos de propiedad intelectual adicionales de terceros.
Yeasen se dedica a la ciencia ética y cree que nuestra investigación debe abordar cuestiones críticas al tiempo que garantiza la seguridad y los estándares éticos.