

Descripción
La D-luciferina es un sustrato común para la luciferasa y se utiliza ampliamente en toda la biotecnología, especialmente en la tecnología de imágenes in vivo. El mecanismo de acción es que la luciferina (el sustrato) se oxida para emitir luz en respuesta al ATP y la luciferasa (ver la figura a continuación). Se produce cuando hay demasiada luciferina. El número cuántico de luz se correlacionó positivamente con la concentración de luciferasa. Los plásmidos que llevan el gen codificador de la luciferasa (Luc) se transfectaron en células y se introdujeron en animales de estudio, como ratas y ratones. In vivo, luego se inyecta fluoresceína y se detectan los cambios en la intensidad de la luz utilizando imágenes de bioluminiscencia (BLI) para monitorear la progresión de la enfermedad o la eficacia del fármaco en tiempo real, etc. La influencia del ATP en el sistema de reacción también se puede utilizar para indicar energía o signos vitales según los cambios en la intensidad de la bioluminiscencia.
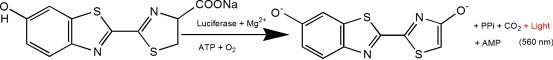
La D-luciferina también se utiliza comúnmente en estudios in vitro, incluidos el análisis de los niveles de luciferasa y ATP; análisis de genes reporteros; secuenciación de alto rendimiento y varias pruebas de contaminación. Actualmente existen tres formas de producto: D-luciferina (ácido libre), sales de D-luciferina (sales de sodio y potasio). La principal diferencia radica en la solubilidad: la primera es soluble en agua y la solubilidad del sistema tampón es débil, excepto soluble en bases débiles como soluciones de NaOH y KOH de baja concentración, soluble en metanol y DMSO; el último es fácilmente soluble en agua o tampón, fácil de usar, solvente no tóxico, especialmente adecuado para experimentos in vivo. Como solución, no existe una diferencia sustancial entre los tres productos en la gran mayoría de aplicaciones.
Características
- Sin radiación, prácticamente inofensivo para los organismos vivos.
- Bioluminiscencia, sin fuente de luz de excitación.
- Es tan sensible que se puede detectar en unos cientos de células.
- Buena penetración, aún se puede detectar una profundidad de tejido de 3-4 cm.
- Alta relación señal-ruido, fuerte señal de fluorescencia y buena antiinterferencia.
Solicitud
- En el experimento de tumorigénesis en ratones desnudos, se observó el crecimiento del tumor sin invasión en tiempo real, sin medición de extirpación del tumor.
- Para probar el efecto de la administración sobre el crecimiento del tumor o la metástasis, el sustrato de fluoresceína puede eliminarse completamente en 3 horas, sin interferir en el experimento farmacológico.
- Se detectó la localización y distribución de células extrañas en animales.
- El gen objetivo o el promotor del gen objetivo se fusiona con el gen de la luciferasa para detectar cambios en la expresión genética durante el tratamiento farmacológico o la progresión de la enfermedad.
- Monitoreo del trasplante de células madre, supervivencia y proliferación; seguimiento de la distribución y migración de células madre in vivo.
Presupuesto
| sinónimo en inglés | (S)-4,5-Dihidro-2-(6-hidroxi-2-benzotiazolil)-4-tiazolcarboxílico ácido sodio sal; D-Luciferina luciérnaga, sal sódica monohidrato; |
| NÚMERO CAS | 103404-75-7 |
| Fórmula | NaClH7N2O3S2·H2O |
| Peso molecular | 320,32 g/mol |
| Apariencia | Polvo amarillo claro |
| Solubilidad | Soluble en agua (100 mg/mL) |
| Pureza (CLAR) | ≥95% |
Componentes
| Componentes No. | Nombre | 40901ES01 | 40901ES02 | 40901ES03 | 40901ES08 | 40901ES10 |
| 40901 | D-luciferina, sal sódica | 100 mg | 500 mg | 1 gramo | 5g | 10 g |
Almacenamiento
El producto debe almacenarse a -15 ℃. ~-25℃ para 1 año.
[1] Guo Y, Guo Y, Chen C, et al. Circ3823 contribuye al crecimiento, la metástasis y la angiogénesis del cáncer colorrectal: participación del eje miR-30c-5p/TCF7. Mol Cancer. 2021;20(1):93. Publicado el 25 de junio de 2021. doi:10.1186/s12943-021-01372-0(IF:27.401)
[2] Wang Z, Yu L, Wang Y, et al. El ajuste dinámico de la atenuación radiativa y no radiactiva de las moléculas AIE refuerza la inmunoterapia y la terapia fototérmica mediadas por imágenes NIR-II. Adv Sci (Weinh). 2022;9(8):e2104793. doi:10.1002/advs.202104793(IF:16.806)
[3] Liu Q, Sheng Z, Cheng C, et al. El anestésico propofol promueve la metástasis tumoral en los pulmones a través de la modulación de la expresión de Src por TRIM21 dependiente de GABA A R. Adv Sci (Weinh). 2021;8(18):e2102079. doi:10.1002/advs.202102079(IF:16.806)
[4] Zhou M, Zhang J, Shen J, et al. La persulfuración de ABI4 unida a sulfuro de hidrógeno controla las respuestas de ABA a través de la transactivación de MAPKKK18 en Arabidopsis. Mol Plant. 2021;14(6):921-936. doi:10.1016/j.molp.2021.03.007(IF:13.164)
[5] Wang J, Wang C, Li Y, et al. Potencial de los exosomas diseñados con péptidos con miR-92b-3p sobreexpresado en la terapia antiangiogénica del cáncer de ovario. Clin Transl Med. 2021;11(5):e425. doi:10.1002/ctm2.425(IF:11.492)
[6] Mao Z, Wei X, Li L, et al. El criptocromo 1 de Arabidopsis controla la fotomorfogénesis mediante la regulación de la deposición de H2A.Z. Plant Cell. 2021;33(6):1961-1979. doi:10.1093/plcell/koab091(IF:11.277)
[7] Xu P, Chen H, Li T, et al. Las interacciones dependientes de la luz azul de CRY1 con las proteínas GID1 y DELLA regulan la señalización de giberelinas y la fotomorfogénesis en Arabidopsis. Plant Cell. 2021;33(7):2375-2394. doi:10.1093/plcell/koab124(IF:11.277)
[8] Lin S, Wen Z, Li S, et al. El lncRNA Neat1 promueve la respuesta inflamatoria de los macrófagos y actúa como un objetivo terapéutico en la osteólisis inducida por partículas de titanio. Acta Biomater. 2022;142:345-360. doi:10.1016/j.actbio.2022.02.007(IF:8.947)
[9] Zhang Z, Zeng D, Zhang W, et al. La modulación de la fosforilación oxidativa aumenta la actividad antineoplásica de la inhibición de la quinasa Aurora mitótica. Cell Death Dis. 2021;12(10):893. Publicado el 30 de septiembre de 2021. doi:10.1038/s41419-021-04190-w(IF:8.469)
[10] Wang L, Tong X, Zhou Z, et al. El ARN circular hsa_circ_0008305 (circPTK2) inhibe la transición epitelial-mesenquimal y la metástasis inducidas por TGF-β al controlar TIF1γ en el cáncer de pulmón de células no pequeñas. Mol Cancer. 2018;17(1):140. Publicado el 27 de septiembre de 2018. doi:10.1186/s12943-018-0889-7(IF:7.776)
[11] Cao X, Xu P, Liu Y, et al.El criptocromo 1 de Arabidopsis promueve el desarrollo estomático a través de la represión de la inhibición de la actividad de unión al ADN de SPEECHLESS por parte de AGB1. J Integr Plant Biol. 2021;63(11):1967-1981. doi:10.1111/jipb.13168(IF:7.061)
[12] Du SS, Li L, Li L, et al. El criptocromo 2 fotoexcitado interactúa directamente con TOE1 y TOE2 en la regulación de la floración. Plant Physiol. 2020;184(1):487-505. doi:10.1104/pp.20.00486(IF:6.902)
[13] Zhou H, Qi Z, Pei P, et al. Nanomicelas biocompatibles para la detección sensible y la terapia fotodinámica del cáncer en etapa temprana. Biomater Sci. 2021;9(18):6227-6235. Publicado el 14 de septiembre de 2021. doi:10.1039/d1bm00847a(IF:6.843)
[14] Zhang L, Zhou J, Yan Y, et al. La nanodispersión sin excipientes de 7-etil-10-hidroxicamptotecina ejerce potentes efectos terapéuticos contra líneas celulares de cáncer de páncreas y xenoinjertos derivados de pacientes. Cancer Lett. 2019;465:36-44. doi:10.1016/j.canlet.2019.08.019(IF:6.508)
[15] Geng Y, Duan H, Xu L, et al. Los modRNA de BMP-2 y VEGF-A en el armazón de colágeno impulsan sinérgicamente la reparación ósea a través de vías osteogénicas y angiogénicas. Commun Biol. 2021;4(1):82. Publicado el 19 de enero de 2021. doi:10.1038/s42003-020-01606-9(IF:6.268)
[16] Qiu J, Peng P, Xin M, et al. Osteólisis periimplantaria inducida por partículas de titanio mediada por ZBTB20 mediante la promoción de respuestas inflamatorias de macrófagos. Biomater Sci. 2020;8(11):3147-3163. doi:10.1039/d0bm00147c(IF:6.183)
[17] Zhou C, Wang G, Jing W, Tan X, Guo H. Propiedades anticancerígenas y mecanismos del polímero de coordinación de plata deshidronorcantaridina de protonación simple en un modelo de cáncer de vejiga. Front Pharmacol. 2021;12:618668. Publicado el 23 de febrero de 2021. doi:10.3389/fphar.2021.618668(IF:5.811)
[18] Sheng Y, Yu H, Pan H, et al. Análisis de todo el genoma de la estructura genética, la expresión y las interacciones proteicas de la familia de genes TIFY del duraznero (Prunus persica). Front Plant Sci. 2022;13:792802. Publicado el 17 de febrero de 2022. doi:10.3389/fpls.2022.792802(IF:5.754)
[19] Jia MZ, Liu LY, Geng C, Jiang J. La activación de las sintasas de ácido 1-aminociclopropano-1-carboxílico establece la densidad estomática y la proporción de agrupamientos en la epidermis de las hojas de Arabidopsis en respuesta a la sequía. Front Plant Sci. 2021;12:758785. Publicado el 6 de diciembre de 2021. doi:10.3389/fpls.2021.758785(IF:5.754)
[20] Xuan Z, Zhao L, Li Z, et al. EPS8L3 promueve la proliferación y metástasis del carcinoma hepatocelular al modular la dimerización e internalización del EGFR. Am J Cancer Res. 2020;10(1):60-77. Publicado el 1 de enero de 2020. (IF:5.177)
[21] Chen G, Fan XY, Zheng XP, Jin YL, Liu Y, Liu SC. Las células madre mesenquimales derivadas del cordón umbilical humano mejoran la resistencia a la insulina a través de la comunicación cruzada mediada por PTEN entre las vías de señalización PI3K/Akt y Erk/MAPKs en los músculos esqueléticos de ratones db/db. Stem Cell Res Ther. 2020;11(1):401. Publicado el 16 de septiembre de 2020. doi:10.1186/s13287-020-01865-7(IF:5.116)
[22] Chen G, Fan XY, Zheng XP, Jin YL, Liu Y, Liu SC. Las células madre mesenquimales derivadas del cordón umbilical humano mejoran la resistencia a la insulina a través de la comunicación cruzada mediada por PTEN entre las vías de señalización PI3K/Akt y Erk/MAPKs en los músculos esqueléticos de ratones db/db. Stem Cell Res Ther. 2020;11(1):401. Publicado el 16 de septiembre de 2020. doi:10.1186/s13287-020-01865-7(IF:5.116)
[23] You D, Du D, Zhao X, Li X, Ying M, Hu X. La enzima málica mitocondrial 2 promueve la metástasis del cáncer de mama mediante la estabilización de HIF-1α en condiciones de hipoxia. Chin J Cancer Res. 2021;33(3):308-322. doi:10.21147/j.issn.1000-9604.2021.03.03(IF:5.087)
[24] Qin L, Zhao R, Chen D, et al. Las células T con receptor de antígeno quimérico dirigidas a PD-L1 suprimen el crecimiento tumoral. Biomark Res. 2020;8:19. Publicado el 3 de junio de 2020. doi:10.1186/s40364-020-00198-0(IF:4.866)
[25] Zuo X, Xiang W, Li K, et al. MdGRF11, un factor regulador del crecimiento, participa en la regulación del tiempo de floración e interactúa con MdTFL1/MdFT1 en la manzana. Plant Sci. 2022;321:111339. doi:10.1016/j.plantsci.2022.111339(IF:4.729)
[26] Yang X, Yu Q, Xu H, Zhou J. La regulación positiva de CD22 por chidamida promueve la funcionalidad de las células T CAR. Sci Rep. 2021;11(1):20637. Publicado el 19 de octubre de 2021. doi:10.1038/s41598-021-00227-4(IF:4.380)
[27] Mei Z, Zhang K, Lam AK, et al. MUC1 como objetivo de la terapia CAR-T en el carcinoma de células escamosas de cabeza y cuello. Cancer Med. 2020;9(2):640-652. doi:10.1002/cam4.2733(IF:3.357)
[28] Zhang Z, Liu L, Cao S, Zhu Y, Mei Q. La administración de genes de TIPE2 inhibe el desarrollo y la metástasis del cáncer de mama a través de respuestas antitumorales mediadas por células T CD8+ y NK. Mol Immunol. 2017;85:230-237. doi:10.1016/j.molimm.2017.03.007(IF:3.236)
[29] Yuan L, Zhou M, Huang D, et al. El resveratrol inhibe la invasión y la metástasis del cáncer de colon mediante la reversión de la transición epitelial-mesenquimal a través de la vía de señalización AKT/GSK-3β/Snail. Mol Med Rep. 2019;20(3):2783-2795. doi:10.3892/mmr.2019.10528(IF:2.952)
[30] Zhang C, Liu X, Chen F, et al. La gliotoxina destruye la función de barrera del epitelio pulmonar al reducir la formación de oligómeros de cofilina para promover la disolución de las fibras de estrés de actina. Microb Pathog. 2018;123:169-176. doi:10.1016/j.micpath.2018.07.007(IF:2.332)
Pago y seguridad
Su información de pago se procesa de forma segura. No almacenamos detalles de la tarjeta de crédito ni tenemos acceso a la información de su tarjeta de crédito.
Consulta
También te puede gustar
Preguntas frecuentes
El producto es solo para fines de investigación y no está destinado a uso terapéutico o diagnóstico en humanos o animales. Los productos y el contenido están protegidos por patentes, marcas comerciales y derechos de autor propiedad de Yeasen Biotechnology. Los símbolos de marca comercial indican el país de origen, no necesariamente el registro en todas las regiones.
Algunas aplicaciones pueden requerir derechos de propiedad intelectual adicionales de terceros.
Yeasen se dedica a la ciencia ética y cree que nuestra investigación debe abordar cuestiones críticas al tiempo que garantiza la seguridad y los estándares éticos.