


Description
Le mélange maître PCR 2×Hieff™ contient de l'ADN polymérase Hieff™ Taq (réf. 10101), des dNTP et d'autres composants requis par la PCR. Le mélange maître est stable pendant 3 mois à 4 °C avec nos stabilisateurs personnalisés. La solution de prémélange est optimisée pour la PCR conventionnelle et prête à l'emploi en ajoutant un modèle d'ADN et des amorces. Les produits PCR peuvent être chargés directement pour l'électrophorèse avec un colorant bleu de bromophénol préchargé. Les produits amplifiés contiennent une protrusion 3'-dA et peuvent être facilement clonés dans le vecteur T. Le mélange maître PCR 2×Hieff™ simplifie la procédure de PCR et réduit la contamination.
Caractéristiques
- Mélange pratique et prêt à l'emploi
- Thermostable : la demi-vie est supérieure à 40 minutes à 95 °C
- Génère des produits PCR avec des surplombs de 3'-dA
Applications
- Amplification PCR de routine de fragments d'ADN
- PCR à haut débit
- Marquage de l'ADN
Caractéristiques
| Fidélité (vs. Taq) | 1 × |
| Démarrage à chaud | Non |
| Surplomb | 3'-A |
| Polymérase | ADN polymérase Taq |
| Format de réaction | SuperMix ou Master Mix |
| Vitesse de réaction | Standard |
| Type de produit | Mélange maître PCR (2×) |
Composants
| Composants N° | Nom | 10102ES03 | 10102ES08 | 10102ES50 | 10102ES60 |
| 10102 | 2×Hieff™ Mélange PCR Master Mix (avec colorant) | 1 ml | 5×1 mL | 50×1 mL | 100×1 mL |
Expédition et stockage
Le 2×Hieff™ Les produits PCR Master Mix doivent être conservés à -15℃ ~ -25℃ depuis 2 ans.
Chiffres
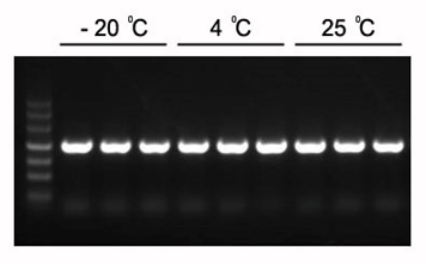
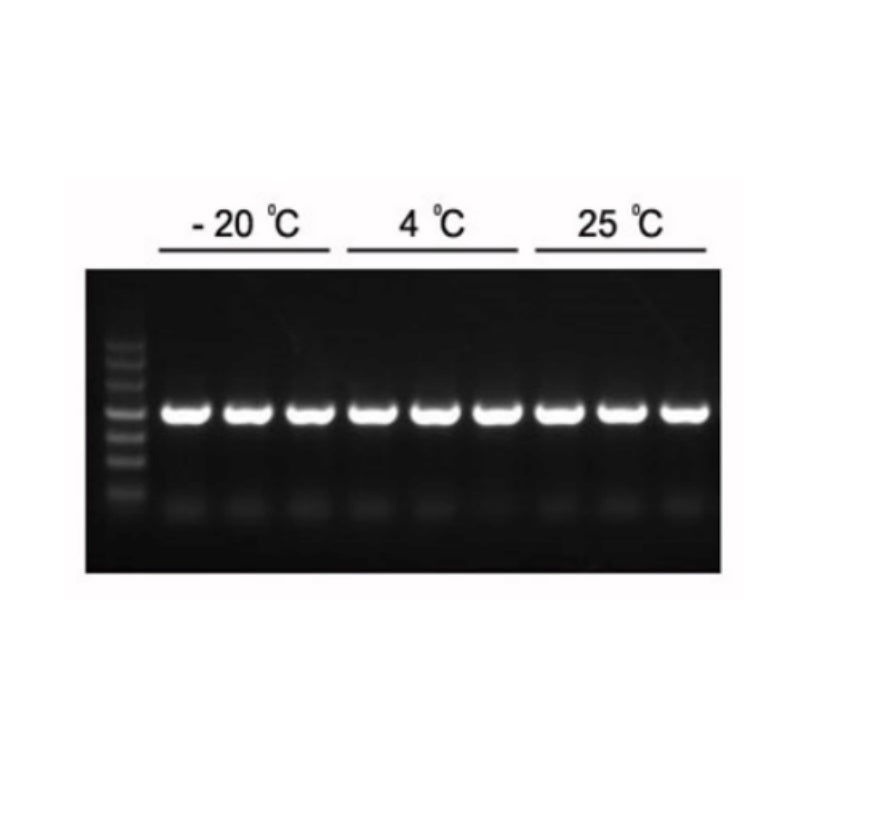
Figure 1. Les produits PCR attendus de 1,2 kb peuvent être amplifiés avec le mélange PCR Master Mix 2×Hieff.
Le Master Mix a été conservé à -20°C pendant 1 an, puis 3 mois à 4°C et 1 mois à 25°C. Modèle : génome d'Arabidopsis. Température de recuit : 60°C. Temps d'extension : 40 sec.
Cité dans « Des capteurs ultrasensibles révèlent le paysage spatiotemporel du métabolisme du lactate dans la physiologie et la maladie »
Cell Metab . 3 janvier 2023 ;35(1):200-211.e9. doi : 10.1016/j.cmet.2022.10.002
[1] Wu LY, Shang GD, Wang FX, et al. Le profilage dynamique de l'état de la chromatine révèle les rôles régulateurs de l'auxine et de la cytokinine dans la régénération des pousses. Dev Cell. 2022;57(4):526-542.e7. doi:10.1016/j.devcel.2021.12.019(IF:12.270)
[2] Li Y, Wang C, Zhang L, Chen B, Mo Y, Zhang J. Claudin-5a est essentiel à la formation fonctionnelle de la barrière hémato-encéphalique et de la barrière hémato-céphalo-rachidienne du poisson zèbre. Fluids Barriers CNS. 2022 ; 19(1) : 40. Publié le 3 juin 2022. doi : 10.1186/s12987-022-00337-9(IF : 7.662)
[3] Luo T, Wang Y, Tang H, et al. Une thérapie génique ciblant NF-κB basée sur l'AAV (rAAV-DMP-miR533) pour les maladies inflammatoires. J Inflamm Res. 2022 ; 15 : 3447-3466. Publié le 14 juin 2022. doi : 10.2147/JIR.S362732(IF : 6.922)
[4] Chen Z, Qi Z, He D, et al. Stratégie de numérisation des ARN circulaires codant pour les peptides dans le cancer colorectal basée sur une analyse bioinformatique et des essais expérimentaux. Front Cell Dev Biol. 2022 ; 9 : 815895. Publié le 25 février 2022. doi : 10.3389/fcell.2021.815895(IF : 6.684)
[5] Yu Y, Fang L. CircRPAP2 régule l'épissage alternatif de PTK2 en se liant à SRSF1 dans le cancer du sein. Cell Death Discov. 2022 ; 8(1) : 152. Publié le 2 avril 2022. doi : 10.1038/s41420-022-00965-y(IF : 5.241)
[6] Zhang J, Liu W, Li G, et al. BCAS2 est impliqué dans l'épissage alternatif et le développement des ovocytes de souris. FASEB J. 2022;36(2):e22128. doi:10.1096/fj.202101279R(IF:5.192)
[7] Xiao W, Li J, Zhang Y, et al. Une souche fongique de Bipolaris bicolor comme bioherbicide potentiel pour la lutte contre l'éleusine indica. Pest Manag Sci. 2022;78(3):1251-1264. doi:10.1002/ps.6742(IF:4.845)
[8] Zhang Y, Yu R, Tang J, et al. Trois gènes de la famille CYP4 du cytochrome P450 régulés par la voie de signalisation CncC induisent une sensibilité phytochimique chez le coléoptère de la farine rouge, Tribolium castaneum. Pest Manag Sci. 2022;78(8):3508-3518. doi:10.1002/ps.6991(IF:4.845)
[9] Zhang X, Yang S, Chen W, et al. ARN circulaire circYPEL2 : un nouveau biomarqueur du cancer du col de l'utérus. Genes (Bâle). 2021 ; 13(1): 38. Publié le 23 décembre 2021. doi : 10.3390/genes13010038(IF : 4.096)
[10] Gu K, Qian D, Qin H, et al. Une nouvelle mutation du gène KCNH2 entraîne une perte de fonction du canal potassique hERG dans le syndrome du QT long 2. Pflugers Arch. 2021 ; 473(2) : 219-229. doi : 10.1007/s00424-021-02518-1(IF : 3.657)
[11] Yang Y, Chu X, Nie M, et al. Une nouvelle délétion à longue portée couvrant STX16 et NPEPL1 provoquant des défauts d'empreinte du locus GNAS découverts chez un patient atteint de pseudohypoparathyroïdie autosomique dominante de type 1B. Endocrine. 2020;69(1):212-219. doi:10.1007/s12020-020-02304-6(IF:3.235)
[12] Ullah H, Arbab S, Khan MIU, et al. Fragment d'ADN mitochondrial circulant sans cellules : un marqueur possible pour la détection précoce de Schistosoma japonicum. Infect Genet Evol. 2021 ; 88 : 104683. doi : 10.1016/j.meegid.2020.104683(IF : 2.773)
[13] Xu L, Chen Y, Shen T, Lin C, Zhang B. Analyse génétique du gène PICK1 dans la maladie d'Alzheimer : une étude pour trouver une nouvelle cible génétique. Front Neurol. 2019 ; 9 : 1169. Publié le 9 janvier 2019. doi : 10.3389/fneur.2018.01169(IF : 2.635)
[14] Ullah H, Qadeer A, Giri BR. Détection d'ADN acellulaire circulant pour diagnostiquer une infection à Schistosoma japonicum. Acta Trop. 2020 ; 211 : 105604. doi : 10.1016/j.actatropica.2020.105604(IF : 2.555)
[15] Yan ZC, Hua HQ, Qi GY, Li YX. Détection et identification précoces des guêpes parasitoïdes Trichogramma Westwood (Hymenoptera : Trichogrammatidae) dans leurs œufs hôtes à l'aide de la réaction en chaîne par polymérase-polymorphisme de longueur des fragments de restriction [publié en ligne avant impression, 24 juin 2022]. J Econ Entomol. 2022;toac095. doi:10.1093/jee/toac095(IF:2.381)
[16] Zhou Z, Yin H, Suye S, Zhu F, Cai H, Fu C. Les changements des cassures double brin de l'ADN et la réparation de l'ADN pendant la formation de la réserve ovarienne chez la souris. Reprod Biol. 2022;22(1):100603. doi:10.1016/j.repbio.2022.100603(IF:2.376)
Paiement et sécurité
Vos informations de paiement sont traitées en toute sécurité. Nous ne stockons pas les détails de la carte de crédit ni accès aux informations de votre carte de crédit.
Enquête
Vous pouvez aussi aimer
FAQ
Le produit est destiné à des fins de recherche uniquement et n'est pas destiné à un usage thérapeutique ou diagnostique chez l'homme ou l'animal. Les produits et le contenu sont protégés par des brevets, des marques déposées et des droits d'auteur appartenant à Yeasen Biotechnology. Les symboles de marque indiquent le pays d'origine, pas nécessairement l'enregistrement dans toutes les régions.
Certaines applications peuvent nécessiter des droits de propriété intellectuelle tiers supplémentaires.
Yeasen se consacre à la science éthique, estimant que nos recherches doivent répondre à des questions cruciales tout en garantissant la sécurité et les normes éthiques.