


Description
Hieff Trans™ Le réactif de transfection liposomale est un réactif de transfection liposomale polyvalent, adapté à la transfection d'ADN, d'ARN et d'oligonucléotides, offrant une efficacité de transfection élevée pour la plupart des cellules eucaryotes. Sa formule unique permet son ajout direct au milieu, et la présence de sérum n'affecte pas l'efficacité de la transfection, ce qui réduit les dommages cellulaires causés par l'élimination du sérum. Il n'est pas nécessaire de retirer le complexe acide nucléique-Hieff Trans™ ni de le remplacer par un milieu frais après la transfection ; il peut également être retiré après 4 à 6 heures.
Hieff Trans™ est fourni sous forme liquide stérile. Habituellement, pour une transfection en plaque 24 puits, environ 1,5 μL à chaque fois, soit 1 mL de Hieff Trans™ peut effectuer environ 660 transfections ; pour une plaque à 6 puits, environ 6 μL à chaque fois, 1 mL de Hieff Trans™ peut effectuer environ 660 transfections. 160 transfections.
Fonctionnalité
- Efficacité exceptionnelle : Performances de transfection supérieures dans une large gamme de lignées cellulaires, y compris la transfection transitoire et la transfection stable
- Grande adaptabilité : Excellente efficacité de transfection dans une variété de lignées cellulaires et niveaux élevés d'expression de protéines recombinantes
- Faible toxicité : les activités des diverses cellules ne sont pratiquement pas affectées par les réactifs de transfection
YEASEN - Fonctionnement simple : efficacité prouvée en présence de sérum — élimine le besoin de changer de milieu après la transfection
- Rentable : effet de transfection compétitif avec des prix plus abordables
Application
- Transfection cellulaire
Spécification
| Formulaire | Liquide |
| Compatible avec le sérum | Oui |
| Type de cellule | Lignées cellulaires établies |
| Type d'échantillon | ADN plasmidique, ARNsi synthétique |
| Technique de transfection | Transfection à base de lipides |
Composants
| Composants n° | Nom | 40802ES02 | 40802ES03 | 40802ES08 |
| 40802 | Hieff Trans™ Réactif de transfection liposomale | 0,5 ml | 1 mL | 5×1 mL |
Expédition et stockage
Le produit est livré avec des blocs réfrigérants et peut être conservé entre 2 et 8 °C pendant un an. Ne pas congeler !
Chiffres

Cité de : Communications Biology volume 5, numéro d'article : 1296 (2022)
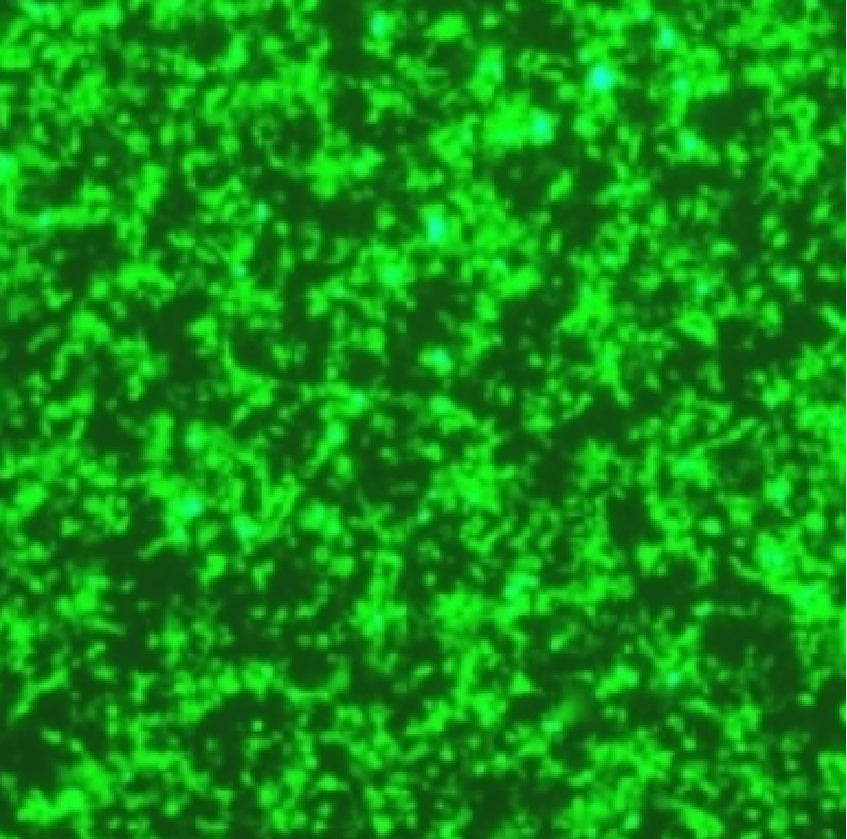
- Démonstration de l'effet de transfection
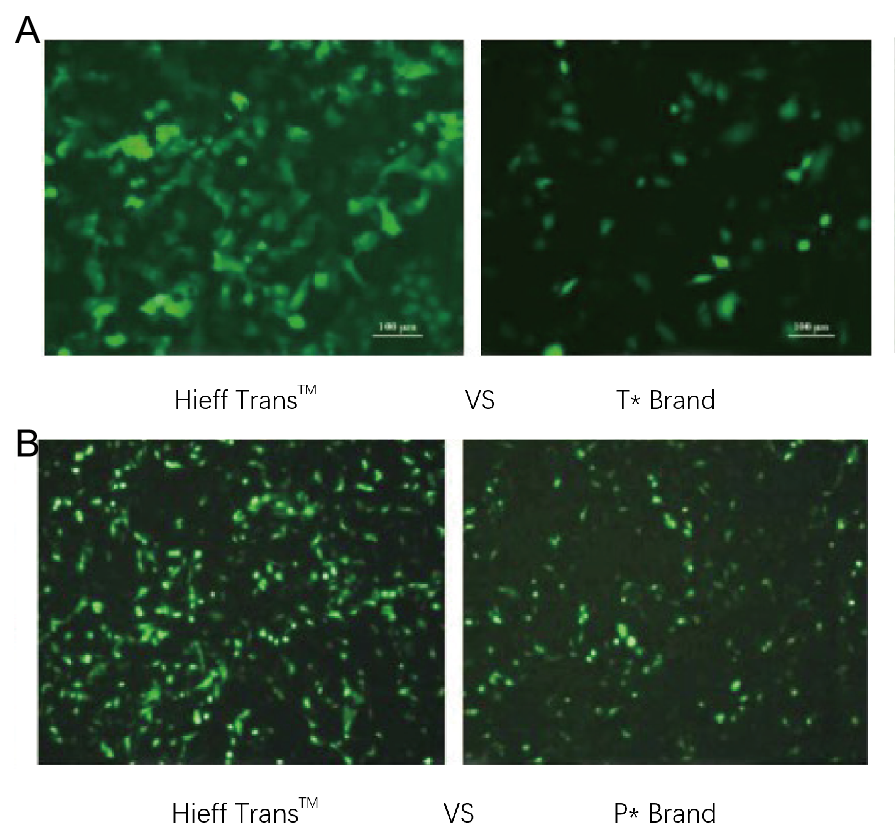
Figure 1. Le réactif de transfection de liposomes Hieff Trans™ surpasse le réactif de transfection de la marque concurrente (marque T* sur la figure 1a, marque P* sur la figure 1b).
Chaque réactif a été utilisé pour transfecter la lignée cellulaire cible (lignée cellulaire Hela dans la figure 1a, lignée cellulaire DF-1 dans la figure 1b) dans un format à 96 puits. L'expression de la GFP a été analysée 48 heures après la transfection. Le réactif de transfection de liposomes Hieff Trans™ a fourni une efficacité de transfection de la GFP supérieure à celle des produits de marques concurrentes.
- Lignées cellulaires validées
| Produit Nom | Hieff TransTMLiposomal Transfection Réactif | |||||
| Validé Cellule Lignes | 293T | Calu 1 | HEK293 | HO1980 | N2A | TS |
| 293F | CHO-K1 | HEK293T | HUVEC | NCI-H1975 | U-87 | |
| 293 pieds | COS-7 | Héla | MCF10A | NIH-3T3 | Véro | |
| 3T3 | DF-1 | Hépatite C | MCF-7 | Neuro-2a | WEHI | |
| A549 | H1299 | Hep3B | MDA-MB-231 | PC-12 | WRL-68 | |
| BV-2 | H520 | Hepa1-6 | MDA-MB-231-LM2-4175 | Râw264.7 | ||
| B50 | HaCaT | HepG2 | MDCK | SGC-7901 | ||
| C2C12 | HCT116 | HK2 | MEF | T47D | … | |
Figure 2. Liste des lignées cellulaires transfectées avec succès avec Hieff Trans™ Réactif de transfection liposomale (en cours de mise à jour continue).

Cité dans « Contrôle optogénétique de la fonction et du métabolisme de l'ARN à l'aide de protéines de liaison à l'ARN photosensibles conçues par génie génétique ». Nat Biotechnol. 3 janvier 2022. doi : 10.1038/s41587-021-01112-1"

Cité de « Structural insight into BRCA1-BARD1 complex recruitment to damaged chromatin. Mol Cell. 2021 Jul 1;81(13):2765-2777.e6. doi: 10.1016/j.molcel.2021.05.010. »
Cité de « L'axe UBQLN2-HSP70 réduit les agrégats de poly-Gly-Ala et atténue les défauts de comportement dans le modèle animal C9ORF72. Neuron. 16 juin 2021 ; 109(12) : 1949-1962.e6. doi : 10.1016/j.neuron.2021.04.023. »
Hieff Trans™ Réactif de transfection liposomale FAQ
(1) Q : Le sérum peut-il être présent lors de la préparation d'un complexe réactif de transfection d'acide nucléique ?
R : La présence de sérum affecte la formation des liposomes. Il est recommandé d'utiliser un milieu sans sérum (généralement un milieu MEM) pour la préparation des complexes réactifs de transfection d'acides nucléiques.
(2) Q : Le réactif de transfection peut-il être congelé ?
R : Non. Ce réactif doit être conservé entre 2 et 8 ℃ et il faut veiller à éviter d'ouvrir le bouchon à plusieurs reprises pendant une longue période, car une ouverture prolongée du bouchon provoquera l'oxydation des liposomes et affectera l'efficacité de la transfection.
(3) Q : À quoi dois-je faire attention lorsque j'utilise le réactif de transfection d'acide nucléique liposomal Hieff Trans™ ?
A : 1) Pendant l'opération de transfection, il est préférable que la confluence cellulaire atteigne 80 à 95 %, et la densité de placage spécifique est déterminée en fonction de la situation des cellules ;
2) L’utilisation d’ADN de haute pureté permet d’obtenir une efficacité de transfection plus élevée ;
3) L'ADN et les réactifs de transfection doivent être dilués avec un milieu sans sérum lors de la préparation des complexes de transfection ;
4) Les antibiotiques ne peuvent pas être ajoutés au milieu pendant la transfection ;
5) La concentration d'ADN et la quantité de réactif liposomal cationique doivent être optimisées dès la première utilisation afin d'obtenir une efficacité de transfection maximale. Le rapport ADN/réactif de transfection est le suivant : il est généralement recommandé d'être dans un rapport de 1:2 à 1:3.
(4) Q : Faut-il l'arrêter après la transfection ?
R : Pas nécessaire. Les complexes liposomiques sont stables pendant 6 heures. Si le milieu cellulaire n'est pas modifié avant la transfection, afin de garantir les nutriments nécessaires à une croissance cellulaire normale, il est nécessaire de changer de milieu après 4 à 6 heures. En revanche, si le milieu a été modifié avant la transfection, il n'est pas nécessaire de le changer après la transfection des liposomes.
(5) Q : À quoi dois-je faire attention si je souhaite améliorer l’efficacité de la transfection ?
A : a : La densité des cellules au moment de la transfection est de 90 à 95 %.
b : Pendant la transfection, utilisez un milieu sans sérum MEM pour les dilutions d'acide nucléique et de liposomes.
c : Le milieu peut être changé 4 à 6 heures après la transfection.
(6) Q : Peut-on réaliser une co-transfection d'ADN et d'ARNsi ? Quel est l'effet ?
R : Une co-transfection peut être réalisée, mais il est recommandé de réaliser une transfection séparée, et la transfection de l'ADN doit être réalisée 6 heures après l'ARNsi. Si les deux opérations sont réalisées simultanément, l'efficacité de la transfection de l'ARNsi sera moindre.
(7) Q : Le réactif de transfection peut-il être utilisé pour la transfection d’emballage lentiviral ?
R : L'emballage lentiviral est possible, mais l'efficacité de l'emballage lentiviral n'est pas nécessairement liée à l'efficacité de la transfection, mais également liée à la sélection des plasmides d'emballage et au rapport entre les plasmides.
(8) Q : Le réactif de transfection d’acide nucléique liposomal Hieff Trans™ peut-il être utilisé pour la transfection de cellules en suspension ?
R : Le réactif de transfection d'acide nucléique liposomique Hieff Trans™ peut être utilisé pour la transfection de cellules en suspension. Voir le protocole pour plus de détails. Nous avons également lancé un réactif de transfection spécifique pour les cellules en suspension (référence : 40805, réactif de transfection d'acide nucléique liposomique pour cellules en suspension).
[1] Liu R, Yang J, Yao J, Zhao Z, He W, Su N, Zhang Z, Zhang C, Zhang Z, Cai H, Zhu L, Zhao Y, Quan S, Chen X, Yang Y. Contrôle optogénétique de la fonction et du métabolisme de l'ARN à l'aide de protéines de liaison à l'ARN photosensibles. Nat Biotechnol. 3 janvier 2022. doi : 10.1038/s41587-021-01112-1. Publication en ligne avant impression. PMID : 34980910. (IF : 54.908)
[2] Zhou J, Chen P, Wang H, Liu H, Li Y, Zhang Y, Wu Y, Paek C, Sun Z, Lei J, Yin L. Variantes de Cas12a conçues pour réduire l'effet hors cible à l'échelle du génome grâce à une reconnaissance rigoureuse des PAM. Mol Ther. 5 janvier 2022 ; 30(1) : 244-255. doi : 10.1016/j.ymthe.2021.10.010. Publication en ligne du 20 octobre 2021. PMID : 34687846 ; PMCID : PMC8753454. (IF : 11.454)
[3] Chen S, Cao X, Zhang J, Wu W, Zhang B, Zhao F. circVAMP3 entraîne la séparation de phase de CAPRIN1 et inhibe le carcinome hépatocellulaire en supprimant la traduction de c-Myc. Adv Sci (Weinh).2022 mars ;9(8) : e2103817. doi : 10.1002/advs.202103817. Publication en ligne du 24 janvier 2022. PMID : 35072355 ; PMCID : PMC8922094. (IF : 16.808)
[4] Zhang Y, Yu X, Sun R, Min J, Tang X, Lin Z, Xie S, Li X, Lu S, Tian Z, Gu C, Teng L, Yang Y. Le facteur d'épissage riche en arginine/sérine 8 favorise la malignité du myélome multiple et les lésions osseuses par l'épissage alternatif de CACYBP et la communication cellulaire basée sur les exosomes. Clin Transl Med. 2022 févr. ; 12(2) : e684. doi : 10.1002/ctm2.684. PMID : 35184390. (IF : 11.492)
[5] Tang X, Deng Z, Ding P, Qiang W, Lu Y, Gao S, Hu Y, Yang Y, Du J, Gu C. Une nouvelle protéine codée par circHNRNPU favorise la progression du myélome multiple en régulant le microenvironnement de la moelle osseuse et l'épissage alternatif. J Exp Clin Cancer Res. 8 mars 2022;41(1):85. est ce que je : 10.1186/s13046-022-02276-7. PMID : 35260179. (SI : 11.161)
[6] Hua Z, Wei R, Guo M, Lin Z, Yu X, Li X, Gu C, Yang Y. YTHDF2 favorise la prolifération des cellules du myélome multiple via l'axe STAT5A/MAP2K2/p-ERK. Oncogene. Mars 2022 ; 41(10) : 1482-1491. doi : 10.1038/s41388-022-02191-3. Publication en ligne du 24 janvier 2022. PMID : 35075244. (IF : 9,867)
[7] Liang Y, Lu Q, Li W, Zhang D, Zhang F, Zou Q, Chen L, Tong Y, Liu M, Wang S, Li W, Ren X, Xu P, Yang Z, Dong S, Zhang B, Huang Y, Li D, Wang H, Yu W. Réactivation du suppresseur de tumeur dans le cancer du sein par commutation d'amplificateur via le réseau NamiRNA. Nucleic Acids Res. 7 septembre 2021 ; 49(15) : 8556-8572. doi : 10.1093/nar/gkab626. PMID : 34329471 ; PMCID : PMC8421228. (SI:16,9)
[8] Dai L, Dai Y, Han J, Huang Y, Wang L, Huang J, Zhou Z. Aperçu structural du recrutement du complexe BRCA1-BARD1 dans la chromatine endommagée. Mol Cell. 1er juillet 2021 ; 81(13) : 2765-2777.e6. doi : 10.1016/j.molcel.2021.05.010. Publication en ligne du 7 juin 2021. PMID : 34102105. (IF : 17,97)
[9] Zhang K, Wang A, Zhong K, Qi S, Wei C, Shu X, Tu WY, Xu W, Xia C, Xiao Y, Chen A, Bai L, Zhang J, Luo B, Wang W, Shen C. L'axe UBQLN2-HSP70 réduit les agrégats de poly-Gly-Ala et atténue les défauts comportementaux dans le modèle animal C9ORF72. Neuron. 16 juin 2021 ; 109(12) : 1949-1962.e6. doi : 10.1016/j.neuron.2021.04.023. Publication en ligne du 14 mai 2021. PMID : 33991504. (IF : 17.17)
[10] Liang Y, Lu Q, Li W, Zhang D, Zhang F, Zou Q, Chen L, Tong Y, Liu M, Wang S, Li W, Ren X, Xu P, Yang Z, Dong S, Zhang B, Huang Y, Li D, Wang H, Yu W. Réactivation du suppresseur de tumeur dans le cancer du sein par commutation d'amplificateur via le réseau NamiRNA. Nucleic Acids Res. 7 septembre 2021 ; 49(15) : 8556-8572. doi : 10.1093/nar/gkab626. PMID : 34329471 ; PMCID : PMC8421228. (IF : 16,9)
[11] Li T, Chen X, Qian Y, Shao J, Li X, Liu S, Zhu L, Zhao Y, Ye H, Yang Y. Dispositif optogénétique synthétique basé sur BRET pour l'expression pulsatile de transgènes permettant l'homéostasie du glucose chez la souris. Nat Commun. 27 janvier 2021 ; 12(1) : 615. doi : 10.1038/s41467-021-20913-1. PMID : 33504786 ; PMCID : PMC7840992. (IF : 14,92)
[12] Pan Y, He X, Li C, Li Y, Li W, Zhang H, Wang Y, Zhou G, Yang J, Li J, Qu J, Wang H, Gao Z, Shen Y, Li T, Hu H, Ma H. L'activité neuronale recrute l'axe CRTC1/CREB pour stimuler l'autophagie dépendante de la transcription afin de maintenir la LTD en phase tardive. Cell Rep. 2021 20 juil. ; 36(3) : 109398. doi : 10.1016/j.celrep.2021.109398. PMID : 34289350. (IF : 9,42)
[13] Liu H, Xing R, Ou Z, Zhao J, Hong G, Zhao TJ, Han Y, Chen Y. Le récepteur couplé aux protéines G GPR17 inhibe le développement du gliome en augmentant la production de ROS médiée par le complexe répressif Polycomb 1. Cell Death Dis. 12 juin 2021 ; 12(6) : 610. doi : 10.1038/s41419-021-03897-0. PMID : 34120140 ; PMCID : PMC8197764. (IF : 8,463)
[14] Fan Y, Wang J, Jin W, Sun Y, Xu Y, Wang Y, Liang X, Su D. CircNR3C2 favorise l'effet suppresseur de tumeur médié par HRD1 via l'épongeage du miR-513a-3p dans le cancer du sein triple négatif. Mol Cancer. 2021 2 février ;20(1):25. doi : 10.1186/s12943-021-01321-x.PMID : 33530981 ; PMCID : PMC7851937. (IF : 27,403)
[15] Gu C, Wang Y, Zhang L, Qiao L, Sun S, Shao M, Tang X, Ding P, Tang C, Cao Y, Zhou Y, Guo M, Wei R, Li N, Xiao Y, Duan J, Yang Y. AHSA1 est une cible thérapeutique prometteuse pour la prolifération cellulaire et la résistance aux inhibiteurs du protéasome dans le myélome multiple. J Exp Clin Cancer Res. 6 janvier 2022 ; 41(1) : 11. doi : 10.1186/s13046-021-02220-1. PMID : 34991674 ; PMCID : PMC8734095. (IF : 11.161)
[16] Luo Q, Wu X, Zhao P, Nan Y, Chang W, Zhu X, Su D, Liu Z. OTUD1 active l'apoptose indépendante et dépendante de la caspase en favorisant la translocation nucléaire d'AIF et la dégradation de MCL1. Adv Sci (Weinh). 8 février 2021 ; 8(8) : 2002874. doi : 10.1002/advs.202002874. PMID : 33898171 ; PMCID : PMC8061361. (IF : 15,84)
[17] Luo Q, Wu X, Nan Y, Chang W, Zhao P, Zhang Y, Su D, Liu Z. TRIM32/USP11 équilibre la stabilité d'ARID1A et le statut oncogène/suppresseur de tumeur du carcinome épidermoïde. Cell Rep. 7 janvier 2020 ; 30(1) : 98-111.e5. doi : 10.1016/j.celrep.2019.12.017. PMID : 31914402. (IF : 9,42)
[18] Sun X, Peng X, Cao Y, Zhou Y, Sun Y. ADNP favorise la différenciation neuronale en modulant la signalisation Wnt/β-caténine. Nat Commun. 12 juin 2020;11(1):2984. est ce que je: 10.1038/s41467-020-16799-0. PMID : 32533114 ; PMCID : PMC7293280. (SI : 14.911)
[19] Yang X, Wang H, Xie E, Tang B, Mu Q, Song Z, Chen J, Wang F, Min J. Le recâblage de la signalisation ERBB3 et ERK confère une résistance à l'inhibition de FGFR1 dans les cancers gastro-intestinaux porteurs d'une mutation ERBB3-E928G. Protein Cell. 2020 déc. ; 11(12) : 915-920. doi : 10.1007/s13238-020-00749-z. PMID : 32632529 ; PMCID : PMC7719122. (IF : 14.872)
[20] Chen, T., Chen, Y., Chen, H. et al. Nanomachine à ADN non lié propulsée par double enzyme pour l'imagerie intracellulaire de microARN faiblement exprimé. Nano Res. 12, 1055–1060 (2019). https://doi.org/10.1007/s12274-019-2344-5 (SI:8.21)
[21] Zhang X, Qi Z, Yin H, Yang G. L'interaction entre les signaux p53 et Ras contrôle la résistance au cisplatine via la régulation de l'apoptose et de l'autophagie médiée par HDAC4 et HIF-1α. Théranostique. 30 janvier 2019 ; 9(4) : 1096-1114. doi : 10.7150/thno.29673. PMID : 30867818 ; PMCID : PMC6401400. (IF : 8.12)
[22] Zou Y, Wang A, Shi M, Chen X, Liu R, Li T, Zhang C, Zhang Z, Zhu L, Ju Z, Loscalzo J, Yang Y, Zhao Y. Analyse des paysages et de la dynamique redox dans les cellules vivantes et in vivo à l'aide de capteurs fluorescents génétiquement codés. Nat Protoc. 2018 oct. ; 13(10) : 2362-2386. doi : 10.1038/s41596-018-0042-5. PMID : 30258175 ; PMCID : PMC6714056. (IF : 13,49)
[23] Zhang K, Zhao X, Chen X, Wei Y, Du W, Wang Y, Liu L, Zhao W, Han Z, Kong D, Zhao Q, Guo Z, Han Z, Liu N, Ma F, Li Z. Effets thérapeutiques améliorés des exosomes dérivés de cellules souches mésenchymateuses avec un hydrogel injectable pour le traitement de l'ischémie des membres postérieurs. ACS Appl Mater Interfaces. 12 septembre 2018 ; 10(36) : 30081-30091. doi : 10.1021/acsami.8b08449. Publication en ligne du 29 août 2018. PMID : 30118197. (IF : 8,09)
[24] Hao H, Hu S, Chen H, Bu D, Zhu L, Xu C, Chu F, Huo X, Tang Y, Sun X, Ding BS, Liu DP, Hu S, Wang M. La perte de CXCR7 endothéliale altère l'homéostasie vasculaire et le remodelage cardiaque après un infarctus du myocarde : implications pour la découverte de médicaments cardiovasculaires. Circulation. 28 mars 2017 ; 135(13) : 1253-1264. doi : 10.1161/CIRCULATIONAHA.116.023027. Publication en ligne du 2 février 2017. PMID : 28154007. (IF : 18.881)
Paiement et sécurité
Vos informations de paiement sont traitées en toute sécurité. Nous ne stockons pas les détails de la carte de crédit ni accès aux informations de votre carte de crédit.
Enquête
Vous pouvez aussi aimer
FAQ
Le produit est destiné à des fins de recherche uniquement et n'est pas destiné à un usage thérapeutique ou diagnostique chez l'homme ou l'animal. Les produits et le contenu sont protégés par des brevets, des marques déposées et des droits d'auteur appartenant à Yeasen Biotechnology. Les symboles de marque indiquent le pays d'origine, pas nécessairement l'enregistrement dans toutes les régions.
Certaines applications peuvent nécessiter des droits de propriété intellectuelle tiers supplémentaires.
Yeasen se consacre à la science éthique, estimant que nos recherches doivent répondre à des questions cruciales tout en garantissant la sécurité et les normes éthiques.