


Descrizione
2×Hieff™ PCR Master Mix contiene Hieff™ Taq DNA Polymerase (Cat#10101), dNTP e altri componenti necessari per la PCR. Il Master Mix è stabile per 3 mesi a 4°C con i nostri stabilizzatori personalizzati. La soluzione premiscelata è ottimizzata per la PCR convenzionale e pronta all'uso aggiungendo il modello di DNA e i primer. I prodotti PCR possono essere caricati direttamente per l'elettroforesi con colorante blu di bromofenolo precaricato. I prodotti amplificati contengono una protrusione di 3 '-dA e possono essere facilmente clonati nel vettore T. Il 2×Hieff™ PCR Master Mix semplifica la procedura PCR e riduce la contaminazione.
Caratteristiche
- Miscela comoda e pronta all'uso
- Termostabile: l'emivita è superiore a 40 minuti a 95°C
- Genera prodotti PCR con sporgenze di 3'-dA
Applicazioni
- Amplificazione PCR di routine dei frammenti di DNA
- PCR ad alta produttività
- Etichettatura del DNA
Specifiche
| Fedeltà (contro Taq) | 1 × |
| Avvio a caldo | NO |
| Sporgenza | 3'-Un |
| Polimerasi | Taq DNA polimerasi |
| Formato di reazione | SuperMix o Master Mix |
| Velocità di reazione | Standard |
| Tipo di prodotto | Miscela Master PCR (2×) |
Componenti
| Componenti n. | Nome | 10102ES03 | 10102ES08 | 10102ES50 | 10102ES60 |
| 10102 | 2×Hieff™ PCR Master Mix (con colorante) | 1 ml | 5×1 ml | 50×1 mL | 100×1 mL |
Spedizione e stoccaggio
Il 2×Hieff™ I prodotti PCR Master Mix devono essere conservati a -15℃ ~ -25℃ per 2 anni.
Cifre
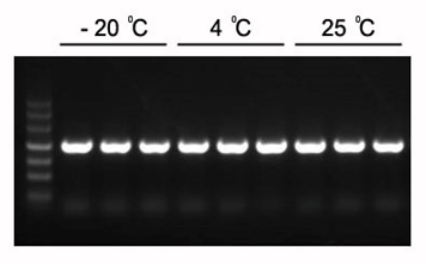
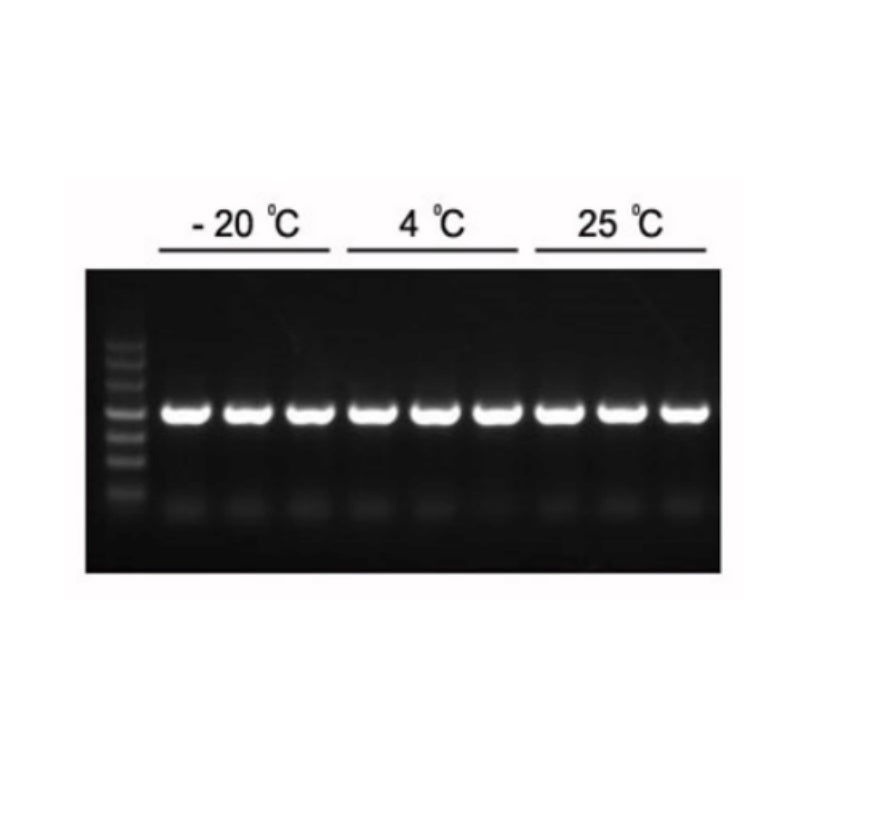
Figura 1. I prodotti PCR previsti da 1,2 kb possono essere amplificati con 2×Hieff PCR Master Mix.
Il Master Mix è stato conservato a -20°C per 1 anno, seguito da altri 3 mesi a 4°C e 1 mese a 25°C. Template: genoma di Arabidopsis. Temperatura di annealing: 60°C. Tempo di estensione: 40 sec.
Citato da "Sensori ultrasensibili rivelano il panorama spaziotemporale del metabolismo del lattato nella fisiologia e nella malattia"
Metabolismo cellulare . 2023 3 gennaio;35(1):200-211.e9. doi: 10.1016/j.cmet.2022.10.002
[1] Wu LY, Shang GD, Wang FX, et al. La profilazione dinamica dello stato della cromatina rivela i ruoli regolatori dell'auxina e della citochinina nella rigenerazione dei germogli. Dev Cell. 2022;57(4):526-542.e7. doi:10.1016/j.devcel.2021.12.019(IF:12.270)
[2] Li Y, Wang C, Zhang L, Chen B, Mo Y, Zhang J. La claudina-5a è essenziale per la formazione funzionale sia della barriera ematoencefalica che della barriera emato-cerebrospinale del pesce zebra. Fluids Barriers CNS. 2022;19(1):40. Pubblicato il 3 giugno 2022. doi:10.1186/s12987-022-00337-9(IF:7.662)
[3] Luo T, Wang Y, Tang H, et al. Una terapia genica mirata a NF-κB basata su AAV (rAAV-DMP-miR533) per le malattie infiammatorie. J Inflamm Res. 2022;15:3447-3466. Pubblicato il 14 giugno 2022. doi:10.2147/JIR.S362732(IF:6.922)
[4] Chen Z, Qi Z, He D, et al. Strategia per la scansione di RNA circolari codificanti peptidi nel cancro del colon-retto basata su analisi bioinformatiche e saggi sperimentali. Front Cell Dev Biol. 2022;9:815895. Pubblicato il 25 febbraio 2022. doi:10.3389/fcell.2021.815895(IF:6.684)
[5] Yu Y, Fang L. CircRPAP2 regola lo splicing alternativo di PTK2 legandosi a SRSF1 nel cancro al seno. Cell Death Discov. 2022;8(1):152. Pubblicato il 2 aprile 2022. doi:10.1038/s41420-022-00965-y(IF:5.241)
[6] Zhang J, Liu W, Li G, et al. BCAS2 è coinvolto nello splicing alternativo e nello sviluppo degli ovociti del topo. FASEB J. 2022;36(2):e22128. doi:10.1096/fj.202101279R(IF:5.192)
[7] Xiao W, Li J, Zhang Y, et al. Un ceppo fungino Bipolaris bicolor come potenziale bioerbicida per il controllo dell'erba oca (Eleusine indica). Pest Manag Sci. 2022;78(3):1251-1264. doi:10.1002/ps.6742(IF:4.845)
[8] Zhang Y, Yu R, Tang J, et al. Tre geni della famiglia del citocromo P450 CYP4 regolati dal percorso di segnalazione CncC mediano la suscettibilità fitochimica nel coleottero della farina rossa, Tribolium castaneum. Pest Manag Sci. 2022;78(8):3508-3518. doi:10.1002/ps.6991(IF:4.845)
[9] Zhang X, Yang S, Chen W, et al. RNA circolare circYPEL2: un nuovo biomarcatore nel cancro cervicale. Genes (Basilea). 2021;13(1):38. Pubblicato il 23 dicembre 2021. doi:10.3390/genes13010038(IF:4.096)
[10] Gu K, Qian D, Qin H, et al. Una nuova mutazione in KCNH2 determina la perdita di funzione del canale del potassio hERG nella sindrome del QT lungo 2. Pflugers Arch. 2021;473(2):219-229. doi:10.1007/s00424-021-02518-1(IF:3.657)
[11] Yang Y, Chu X, Nie M, et al. Una nuova delezione a lungo raggio che si estende su STX16 e NPEPL1 causando difetti di imprinting del locus GNAS scoperti in un paziente con pseudoipoparatiroidismo autosomico dominante di tipo 1B. Endocrino. 2020;69(1):212-219. doi:10.1007/s12020-020-02304-6(IF:3.235)
[12] Ullah H, Arbab S, Khan MIU, et al. Frammento di DNA mitocondriale libero circolante: un possibile marcatore per la rilevazione precoce di Schistosoma japonicum. Infect Genet Evol. 2021;88:104683. doi:10.1016/j.meegid.2020.104683(IF:2.773)
[13] Xu L, Chen Y, Shen T, Lin C, Zhang B. Analisi genetica del gene PICK1 nella malattia di Alzheimer: uno studio per trovare un nuovo bersaglio genetico. Front Neurol. 2019;9:1169. Pubblicato il 9 gennaio 2019. doi:10.3389/fneur.2018.01169(IF:2.635)
[14] Ullah H, Qadeer A, Giri BR. Rilevamento del DNA libero circolante per diagnosticare l'infezione da Schistosoma japonicum. Acta Trop. 2020;211:105604. doi:10.1016/j.actatropica.2020.105604(IF:2.555)
[15] Yan ZC, Hua HQ, Qi GY, Li YX. Rilevamento precoce e identificazione delle vespe parassite Trichogramma Westwood (Hymenoptera: Trichogrammatidae) nelle loro uova ospiti utilizzando il polimorfismo della lunghezza del frammento di restrizione della reazione a catena della polimerasi [pubblicato online prima della stampa, 24 giugno 2022]. J Econ Entomol. 2022;toac095. doi:10.1093/jee/toac095(IF:2.381)
[16] Zhou Z, Yin H, Suye S, Zhu F, Cai H, Fu C. I cambiamenti delle rotture del doppio filamento del DNA e la riparazione del DNA durante la formazione della riserva ovarica nei topi. Reprod Biol. 2022;22(1):100603. doi:10.1016/j.repbio.2022.100603(IF:2.376)
Pagamento e sicurezza
Le informazioni di pagamento vengono elaborate in modo sicuro. Non archiviamo i dettagli della carta di credito né abbiamo accesso alle informazioni sulla tua carta di credito.
Indagine
Potrebbe piacerti anche
FAQ
Il prodotto è solo per scopi di ricerca e non è destinato all'uso terapeutico o diagnostico su esseri umani o animali. Prodotti e contenuti sono protetti da brevetti, marchi e copyright di proprietà di Yeasen Biotechnology. I simboli dei marchi indicano il paese di origine, non necessariamente la registrazione in tutte le regioni.
Alcune applicazioni potrebbero richiedere ulteriori diritti di proprietà intellettuale di terze parti.
Yeasen è un sostenitore della scienza etica, convinto che la nostra ricerca debba affrontare questioni critiche garantendo al contempo sicurezza e standard etici.