

描述
GMyc-PCR支原體檢測試劑盒主要利用PCR方法檢測各種生物材料(如細胞培養、實驗動物分泌物、動物血清等)的支原體感染。它結合了多種優點:靈敏、特異、快速,並且可以直接用細胞培養上清液檢測。本產品採用PCR方法檢測培養細胞等生物材料中的支原體。所用引子依支原體16S-23S rRNA序列保守區設計,只特異性擴增支原體DNA,檢測敏感度高、特異性強。 PCR擴增及電泳分析僅需數小時,操作方便簡單。
細胞培養是生命科學研究中常見的實驗。與其他常用的實驗方法不同,細胞培養是一個動態的連續過程,細胞通常會對操作錯誤或污染物作出反應,而這些錯誤或污染物常常表現出異常的細胞狀態或培養基外觀。若受到支原體污染,細胞形態無明顯改變,容易被忽略。往往直到污染非常嚴重的時候才被發現。被污染的細胞膜上可能有上百個支原體,這些支原體會爭奪營養物質並釋放有毒代謝產物,嚴重影響實驗結果。
研究表明,至少有20種支原體可以污染細胞,其中最常見的有:口腔支原體(M. orale)、精氨酸支原體(精胺酸小體)、豬鼻支原體(豬鼻支原體)、發酵支原體(發酵支原體)、人型支原體(人類型支原體)、唾液支原體(唾液分枝桿菌)、肺支原體(M. 肺支原體)和梨支原體(M. pirum)。培養細胞的支原體污染率為4%~92%。污染源包括工作環境、操作者本身(某些支原體是人體的正常菌群)、培養基、血清、細胞交叉污染、實驗器材、以及用來製備細胞的原始組織或器官的污染。
確定細胞培養過程中問題的根本原因是一項困難且耗時的任務,任何突然的變化都應該懷疑,良好的測試實踐和定期的支原體污染檢測是必要的。支原體的檢測方法很多,如直接培養法、DNA螢光染色法、ELISA法、PCR法等。
特徵
- 所用引子依支原體16S-23S rRNA序列的保守區設計
- 僅特異性擴增支原體DNA
- 優異的敏感性和特異性
- 可偵測到 20 多種支原體
應用
- 支原體檢測
成分
| 零件編號 | 姓名 | 40601ES10 (10 次測定) | 40601ES20 (20 次檢測) |
| 40601-A | GMyc-1st PCR 混合物 | 250 微升 | 2×250 µL |
| 40601-B | GMyc-2nd PCR 混合物 | 250 微升 | 2×250 µL |
| 40601-C | 陽性對照模板 N | 20 微升 | 20 微升 |
【注意事項】1.長期不使用時,可冷凍保存於-85~-65℃。
2. PCR反應極為敏感。為了防止假陽性,在最後加入樣本時會加入陽性對照。
貯存
本產品在-25~-15℃可保存18個月。若長時間不使用,請遠離光照。
數位
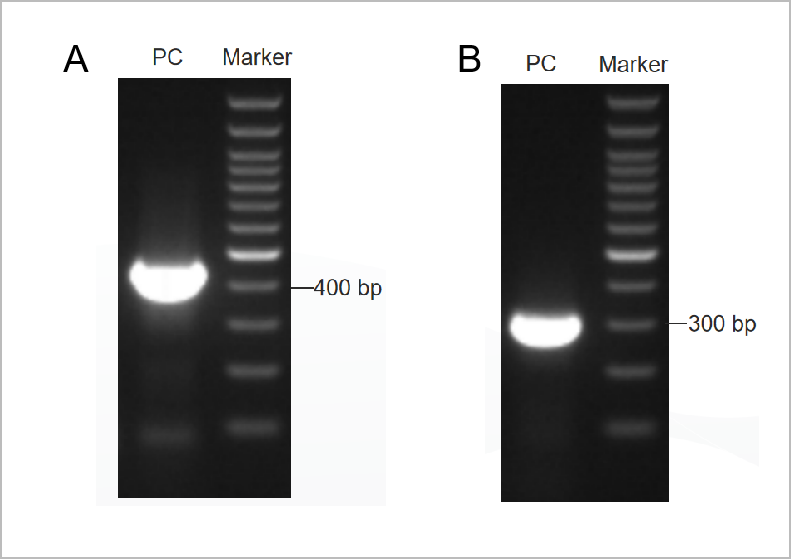
圖1. PCR支原體檢測試劑盒電泳圖譜結果。
第一輪電泳圖如圖1A所示,第二輪電泳圖如圖1B所示。
[1] 饒曉松, 叢曉霞, 高曉康, 等. AMPK 介導的磷酸化增強 TBC1D17 的自身抑制,從而促進 Rab5 依賴的葡萄糖吸收。細胞死亡有所不同。 2021;28(12):3214-3234。 doi:10.1038/s41418-021-00809-9(影響因子:15.828)
[2] 郭鋒, 李玲, 李菁, 等.小鼠早期胚胎和胚胎幹細胞的單細胞多組學定序。細胞研究。 2017;27(8):967-988。 doi:10.1038/cr.2017.82(影響因子:15.606)
[3] 郝燕, 何斌, 吳玲, 等. p85β 的核易位促進 PIK3CA 螺旋結構域突變癌症的腫瘤發生。國家通訊社2022;13(1):1974。出版日期:2022 年 4 月 13 日。
[4] 舒曉, 劉梅, 陸哲, 等.全基因組圖譜顯示脫氧尿苷在人類著絲粒 DNA 中富集。天然化學生物學。 2018;14(7):680-687。 doi:10.1038/s41589-018-0065-9(影響因子:13.843)
[5] 李曉玲, 熊小玲, 王凱, 等.轉錄組範圍的映射揭示了可逆的和動態的N(1)-甲基腺苷甲基化組。天然化學生物學。 2016;12(5):311-316。 doi:10.1038/nchembio.2040(影響因子:12.709)
[6] 孫琳,楊曉玲,黃曉玲,等.脂肪酸的 2-羥基化透過 YAP 轉錄軸抑制大腸直腸腫瘤發生和轉移。癌症研究。 2021;81(2):289-302。 doi:10.1158/0008-5472.CAN-20-1517(影響因子:12.701)
[7] 孫哲, 張哲, 王慶, 劉建玲. CTPS1 和 ATR 的合併失活對 MYC 過度表現的癌細胞具有合成致死性。癌症研究。 2022;82(6):1013-1024。 doi:10.1158/0008-5472.CAN-21-1707(影響因子:12.701)
[8] 宋江, 莊毅, 朱聰, 等.人類 PUS10 在 miRNA 加工和 tRNA 假尿苷化中的不同作用。天然化學生物學。 2020;16(2):160-169。 doi:10.1038/s41589-019-0420-5(影響因子:12.154)
[9] 何斌, 潘華, 鄭鋒, 等.長鏈非編碼RNA LINC00930促進PFKFB3介導的鼻咽癌腫瘤糖解作用和細胞增殖。 J Exp Clin Cancer Res. 2022;41(1):77。出版日期:2022 年 2 月 24 日。
[10] 唐斌,劉斌華,劉志勇,羅明陽,石曉華,龐德偉.具有緻密兩親性兩性離子塗層的量子點。 ACS 應用材料接口。 2022;14(24):28097-28104。 doi:10.1021/acsami.2c04438(影響因子:9.229)
[11] 黃聰, 張哲, 陳玲, 等.Src 的 N 端和 C 端結構域內的乙醯化調節 STAT3 介導的腫瘤發生的獨特作用。癌症研究。 2018;78(11):2825-2838。 doi:10.1158/0008-5472.CAN-17-2314(影響因子:9.130)
[12] 吳曉, 餘梅, 張哲, 等. DDB2 透過 PCNA 獨立的 CDT2 降解來調節 DNA 複製。細胞生物科學。 2021;11(1):34。出版日期:2021 年 2 月 8 日。
[13] 王建軍, 張燕, 劉曉玲, 劉紅. 依腫瘤抗藥性亞群可達性優化自適應療法。癌症(巴塞爾)。 2021;13(21):5262。 2021 年 10 月 20 日出版。
[14] 馮偉, 劉榮, 謝曉, 等. α-微管蛋白的 SUMO 化是一種調節微管動力學的新型修飾。 J Mol Cell Biol. 2021;13(2):91-103。 doi:10.1093/jmcb/mjaa076(影響因子:6.216)
[15] Yu M, Hu X, Yan J, Wang Y, Lu F, Chang J. RIOK2 抑制劑 NSC139021 透過誘導 Skp2 介導的細胞週期停滯和細胞凋亡對膠質母細胞瘤發揮抗腫瘤作用。生物醫藥。 2021;9(9):1244。出版日期:2021 年 9 月 17 日。
[16] 任紹軍, 蔡穎, 胡珊, 等.小檁鹼可透過調節 c-myc/CD47 軸在瀰漫性大 B 細胞淋巴瘤中發揮抗腫瘤活性。生物化學藥理學。 2021;188:114576。 doi:10.1016/j.bcp.2021.114576(影響因子:5.858)
[17] 溫鋒, 孫小玲, 孫晨, 等. TAGLN 在前列腺癌細胞中因 TRAF6 介導的蛋白酶體降解而下調。 Mol Cancer Res。 2021;19(7):1113-1122。 doi:10.1158/1541-7786.MCR-20-0513(影響因子:5.852)
[18] 唐斌, 孫永忠, 張志玲, 等.鞘磷脂隔離的膽固醇結構域會招募福馬林結合蛋白 17,以收縮流感病毒進入時的網格蛋白包被小窩。 J Virol. 2022;96(5):e0181321。 doi:10.1128/JVI.01813-21(影響因子:5.103)
[19] 胡菁, 任偉, 邱偉, 等.從患有JAK2 突變的僵直性脊椎炎患者產生誘導多能幹細胞系 (XDCMHi001-A)。幹細胞研究。 2020;45:101788。 doi:10.1016/j.scr.2020.101788(影響因子:4.495)
[20] Xiao S, Yao X, Ye J, Tian X, Yin Z, Zhou L. 表觀遺傳修飾促進脯胺酸合成酶 PYCR1 在胃癌中的異常表現 [2022 年 5 月 30 日線上發表]。 Biochim Biophys Acta Gene Regul Mech. 2022;1865(6):194829。 doi:10.1016/j.bbagrm.2022.194829(影響因子:4.490)
[21] 王菁, 張燕, 劉曉玲, 劉紅. 在資訊不完整的情況下, 固定週期治療對腫瘤系統是否有效?癌症管理研究。 2021;13:8915-8928。出版日期:2021 年 11 月 30 日。
[22] Yang X, Ren S, Rehman ZU 等.牛 (Bos taurus) 和山羊 (Capra hircus) 的 TANK 結合激酶 1 (TBK1) 的分子表徵、表達和功能鑑定。 Dev Comp Immunol。 2022;133:104444。 doi:10.1016/j.dci.2022.104444(影響因子:3.636)
[23] 鄭丹, 常曉玲, 劉艷, 等. 2-甲氧基-5((3,4,5-三甲氧基苯基)硒基)苯酚透過下調乳癌細胞系中的 MDM2 逆轉 EGF 誘導的細胞遷移和侵襲。癌症生物治療。 2019;20(4):513-523。 doi:10.1080/15384047.2018.1537578(影響因子:3.373)
[24] 徐鋒, 張勝, 劉哲, 等. TEX9 和 eIF3b 發揮協同作用,促進食道鱗狀細胞癌的進展。 BMC 癌症。 2019;19(1):875。出版日期:2019 年 9 月 3 日。
[25] 潘華, 孫玲, 王偉, 等.血清長鏈非編碼 RNA LOC553103 作為常見人類癌症的非特異性診斷和預後生物標記。 Clin Chim Acta. 2020;508:69-76。 doi:10.1016/j.cca.2020.05.017(影響因子:2.615)
[26] 李娜, 林紹明, 李燕, 孫建, 張玲, 陳梅.誘導性多能幹細胞系 (GZHMCi004-A) 源自於 FGFR3 基因雜合 G380R 突變而導致軟骨發育不全的胎兒。幹細胞研究。 2021;53:102322。 doi:10.1016/j.scr.2021.102322(影響因子:2.020)
[27] 羅倩,韋聰,龍毅,等.透過 iCRISPR/Cas9 系統產生 ELTD1 敲除人類胚胎幹細胞株。幹細胞研究。 2021;53:102350。 doi:10.1016/j.scr.2021.102350(影響因子:2.020)
[28] 劉永強, 凌天偉, 王紅艷, 楊永華, 宋文江, 王TC.從患有 DGKG 突變的癲癇患者中生成無整合誘導多能幹細胞系 (LZUSHI001-A)。幹細胞研究。 2022;61:102768。 doi:10.1016/j.scr.2022.102768(影響因子:2.020)
[29] 陳梅,林紹波,李娜,李燕,李毅,張玲. 誘導性多能幹細胞系(GZHMCi003-A)來自 RUNX2 基因外顯子 3 雜合缺失導致顱骨發育不良的胎兒。幹細胞研究。 2021;51:102166。 doi:10.1016/j.scr.2021.102166(影響因子:2.020)
[30] 徐穎, 王鑫, 邱婷, 等.產生誘導性多能幹細胞系 (FDCHI007-A),源自攜帶雜合 c.545C 的 31 型發育性和癲癇性腦病變患者 > DNM1 基因發生突變。幹細胞研究。 2022;60:102709。 doi:10.1016/j.scr.2022.102709(影響因子:2.020)
[31] 範婷,何建,王艷,餘建,孫偉. 從一名7歲中樞性性早熟女孩體內建立誘導性多能幹細胞系(FDCHi006-A)。幹細胞研究。 2021;56:102542。 doi:10.1016/j.scr.2021.102542(影響因子:2.020)
[32] Gong X, Zheng Z, Yang T, Zheng H, Xiao X, Jia N. 從具有 ARX 基因半合子 R330L 突變的 1 型早期嬰兒癲癇性腦病 (EIEE1) 患者中生成同源基因校正的 iPSC 系 (OGHFUi001-A-1)。幹細胞研究。 2022;60:102693。 doi:10.1016/j.scr.2022.102693(影響因子:2.020)
[33]賈娜, 龔曉, 陳菁, 等.從患有 ARX 突變的 1 型早期嬰兒癲癇性腦病變中生成誘導性多能幹細胞系 (OGHFUi001-A)。幹細胞研究。 2021;53:102367。 doi:10.1016/j.scr.2021.102367(影響因子:2.020)
[34] 朱偉, 週艷, 王倩, 等.從攜帶 PSEN1 基因 E363Q 突變的患者的皮膚纖維母細胞生成人類誘導多能幹細胞 (iPSC) 系。幹細胞研究。 2022;61:102769。 doi:10.1016/j.scr.2022.102769(影響因子:2.020)
[35] 羅非, 龍凱, 李曉玲, 等. LRRC8A 缺乏會減輕 3T3-L1 細胞缺氧引起的壞死。 Biosci 生物技術 生物化學。 2020;84(6):1139-1145。 doi:10.1080/09168451.2020.1730689(影響因子:1.516)
付款和安全
您的付款信息已安全處理。我們無法存儲信用卡詳細信息,也無法訪問您的信用卡信息。
詢問
你也可能喜歡
常問問題
本產品僅用於研究目的,不用於人類或動物的治療或診斷用途。產品和內容受以下公司的專利、商標和版權保護:
某些應用程式可能需要額外的第三方智慧財產權。