


描述
希夫傳輸™ 脂質體轉染試劑是一種多功能的脂質體轉染試劑,適用於DNA、RNA和寡核苷酸的轉染,對大多數真核細胞具有較高的轉染效率。其獨特的配方使其可直接加入培養基中,血清的存在不會影響轉染效率,減少了因去除血清對細胞的傷害。轉染後無需移除核酸-Hieff Trans™複合物或更換新鮮培養基,也可在4-6小時後移除。
希夫傳輸™ 以無菌液體形式供應。通常,對於24孔板轉染,每次約1.5 μL,1 mL Hieff Trans™ 可進行約660次轉染; 6孔板,每次約6 μL,1 mL Hieff Trans™可進行約660次轉染。 160 次轉染。
特徵
- 卓越的效率:在廣泛的細胞系中具有卓越的轉染性能,包括瞬時轉染和穩定轉染
- 適應性廣:在多種細胞系中均具有優異的轉染效率,且重組蛋白表現量高
- 低毒性:轉染試劑幾乎不影響各種細胞的活動
YEASEN - 操作簡單:在血清存在下已證實有效-轉染後無需更換培養基
- 成本效益:具競爭力的轉染效果,價格較實惠
應用
- 細胞轉染
規格
| 形式 | 液體 |
| 相容精華素 | 是的 |
| 單元格類型 | 已建立細胞系 |
| 樣品類型 | 質粒 DNA、合成 siRNA |
| 轉染技術 | 脂質轉染 |
成分
| 零件編號 | 姓名 | 40802ES02 | 40802ES03 | 40802ES08 |
| 40802 | 希夫譯™ 脂質體轉染試劑 | 0.5 毫升 | 1 毫升 | 5×1 毫升 |
運輸和儲存
產品附帶冰袋運輸,可在2-8ºC下保存一年。不要凍結!
數位

引自: 通訊生物學第 5 卷,文章編號:1296 (2022)
- 轉染效果驗證
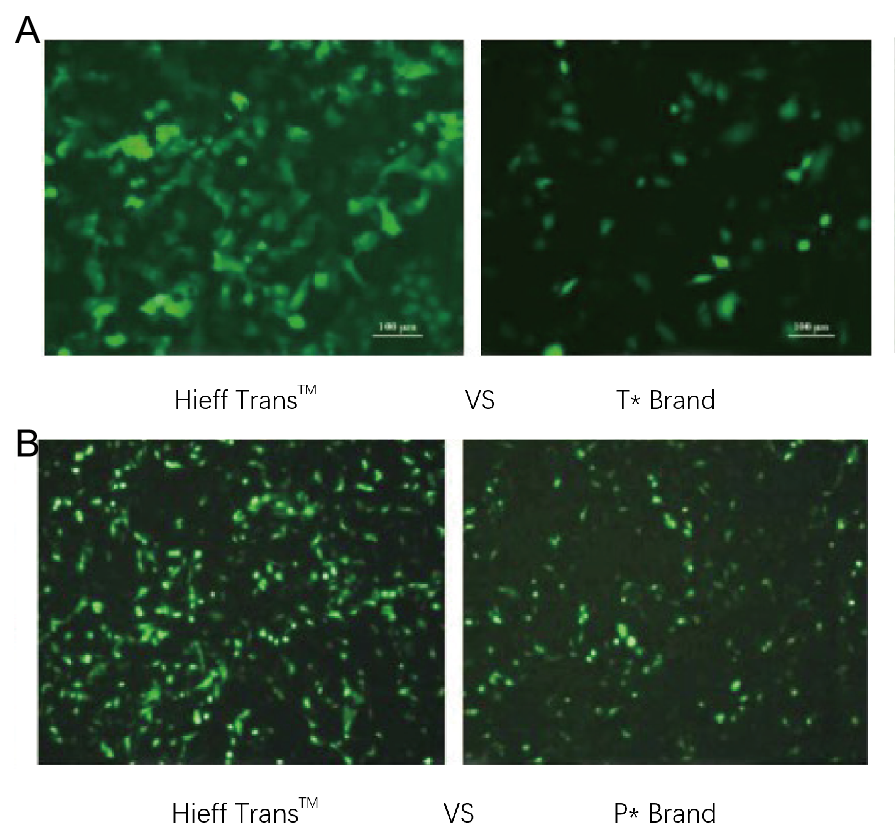
圖1. Hieff Trans™脂質體轉染試劑的性能優於競爭品牌的轉染試劑(圖1a為T*品牌,圖1b為P*品牌)。
每種試劑均以96孔板形式轉染目標細胞系(圖1a中的Hela細胞系,圖1b中的DF-1細胞系)。轉染後48小時分析GFP表現。 Hieff Trans™脂質體轉染試劑比競爭品牌產品提供更高的GFP轉染效率。
- 已驗證的細胞系
| 產品 姓名 | 希夫 透皮脂質體 轉染 試劑 | |||||
| 已驗證 細胞 線條 | 293T | 卡盧 1 | HEK293 | HO1980 | N2A | TS |
| 293F | CHO-K1 | HEK293T | 臍靜脈內皮細胞 | NCI-H1975 | U-87 | |
| 293 英尺 | COS-7 | 希拉 | MCF10A | NIH-3T3 | 維羅 | |
| 3T3 | 東風-1 | Hep2C | MCF-7 | 神經-2a | 威海 | |
| A549 | H1299 | Hep3B | MDA-MB-231 | PC-12 | WRL-68 | |
| BV-2 | H520 | 肝素1-6 | MDA-MB-231-LM2-4175 | 鐳w264.7 | ||
| B50 | 肝細胞癌 | 肝癌 | 糖尿病腎病變 | SGC-7901 | ||
| C2C12 | HCT116 | HK2 | 中東呼吸聯合會 | T47D | … | |
圖 2. 成功轉染 Hieff Trans™ 的細胞系列表 脂質體轉染試劑(持續更新中)。

引自「利用工程化光開關 RNA 結合蛋白進行 RNA 功能和代謝的光遺傳學控制。Nat Biotechnol。2022 年 1 月 3 日。doi:10.1038/s41587-021-01112-1"

引自“BRCA1-BARD1 複合物募集至受損染色質的結構洞察。Mol Cell。2021 年 7 月 1 日;81(13):2765-2777.e6。doi:10.1016/j.molcel.2021.05.010。”
引自「UBQLN2-HSP70 軸減少聚-Gly-Ala 聚集體並減輕 C9ORF72 動物模型中的行為缺陷。神經元。2021 年 6 月 16 日;109(12):1949-1962.e6.doi: 10.101/j.j020.10.1020.
希夫傳輸™ 脂質體轉染試劑 常問問題
(1)問:製備核酸轉染試劑複合物時可以存在血清嗎?
答:血清的存在會影響脂質體的形成。建議在製備核酸轉染試劑複合物時使用無血清培養基(通常為MEM培養基)。
(2)問:轉染試劑可以冷凍嗎?
答:不行。
(3)問:使用Hieff Trans™脂質體核酸轉染試劑時應注意什麼?
A:1)轉染操作時,細胞匯合度達80%-95%為佳,具體接種密度依細胞狀況決定;
2)使用高純度的DNA有助於獲得更高的轉染效率;
3)製備轉染複合物時需以無血清培養基稀釋DNA及轉染試劑;
4)轉染時培養基中不能添加抗生素;
5)首次使用時應優化DNA濃度和陽離子脂質體試劑的用量,以獲得最大的轉染效率。 DNA與轉染試劑的比例為 一般建議為1:2-1:3。
(4)轉染後需要終止嗎?
答:不需要。脂質體複合物可穩定保存 6 小時。如果轉染前沒有更換細胞培養基,為了確保細胞正常生長所需的營養,需在4~6小時後再換成新培養基。然而,如果轉染前已更換培養基,則脂質體轉染後無需更換培養基。
(5)問:提高轉染效率該注意什麼?
答:a:轉染時細胞密度為90%-95%。
b:轉染時使用MEM無血清培養基進行核酸和脂質體稀釋。
c:轉染後4-6小時可換液。
(6)問:DNA和siRNA可以共轉染嗎?效果如何?
A:可共轉染,但建議單獨轉染,且DNA轉染應在siRNA轉染6小時後進行。若兩者聯合操作,siRNA的轉染效率會較差。
(7)此轉染試劑可以用於慢病毒包裝轉染嗎?
A:慢病毒包裝是可以的,但是慢病毒包裝的效率和轉染的效率沒有必然聯繫,還和包裝質粒的選擇以及質粒之間的配比有關。
(8)問:Hieff Trans™脂質體核酸轉染試劑可以用於懸浮細胞的轉染嗎?
答:Hieff Trans™脂質體核酸轉染試劑可用於懸浮細胞轉染,詳情請參考實驗方案。此外,我們也推出了專門針對懸浮細胞的轉染試劑(Cat No.40805,懸浮細胞脂質體核酸轉染試劑)。
[1] Liu R, Yang J, Yao J, Zhao Z, He W, Su N, Zhang Z, Zhang C, Zhang Z, Cai H, Zhu L, Zhao Y, Quan S, Chen X, Yang Y. 利用工程化光開關 RNA 結合蛋白進行光遺傳學控制 RNA 功能和代謝。天然生物技術。 2022 年 1 月 3 日。 Epub 優先印刷。 PMID: 34980910.(影響因子:54.908)
[2] Zhou J, Chen P, Wang H, Liu H, Li Y, Zhang Y, Wu Y, Paek C, Sun Z, Lei J, Yin L. Cas12a 變異體設計用於透過嚴格的 PAM 識別降低全基因組脫靶效應。分子治療學2022年1月5日;30(1):244-255。 doi: 10.1016/j.ymthe.2021.10.010。 Epub 2021 年 10 月 20 日。 PMCID:PMC8753454。 (影響因子:11.454)
[3] Chen S, Cao X, Zhang J, Wu W, Zhang B, Zhao F. circVAMP3 透過抑制 c-Myc 翻譯驅動 CAPRIN1 相分離並抑制肝細胞癌。 Adv Sci(Weinh)。2022 年 3 月;9(8):e2103817。 doi: 10.1002/advs.202103817。 Epub 2022 年 1 月 24 日。 PMCID:PMC8922094。 (影響因子:16.808)
[4] 張艷,餘曉玲,孫榮,閔建軍,唐曉玲,林哲,謝勝,李曉玲,陸勝,田哲,顧晨,滕玲,楊燕. 剪接因子精氨酸/富含絲氨酸 8 通過 CACYBP 的可變剪接和基於外泌體的細胞通訊促進多發性骨髓瘤惡性腫瘤和骨病變。臨床翻譯醫學。 2022 年 2 月;12(2):e684。 doi: 10.1002/ctm2.684。 PMID: 35184390.(影響因子:11.492)
[5] Tang X, Deng Z, Ding P, Qiang W, Lu Y, Gao S, Hu Y, Yang Y, Du J, Gu C. circHNRNPU 編碼的一種新蛋白透過調節骨髓微環境和可變剪接促進多發性骨髓瘤進展。 J Exp Clin Cancer Res. 2022年3月8日;41(1):85。 doi: 10.1186/s13046-022-02276-7。 PMID: 35260179. (影響因子:11.161)
[6] Hua Z, Wei R, Guo M, Lin Z, Yu X, Li X, Gu C, Yang Y. YTHDF2 透過 STAT5A/MAP2K2/p-ERK 軸促進多發性骨髓瘤細胞增生。致癌基因。 2022年3月;41(10):1482-1491。 doi: 10.1038/s41388-022-02191-3。 Epub 2022 年 1 月 24 日。
[7] Liang Y, Lu Q, Li W, Zhang D, Zhang F, Zou Q, Chen L, Tong Y, Liu M, Wang S, Li W, Ren X, Xu P, Yang Z, Dong S, Zhang B, Huang Y, Li D, Wang H, Yu W. 透過 NamiRNA 網路增強子轉換重新活化乳癌中的腫瘤抑制基因。核酸研究。 2021年9月7日;49(15):8556-8572。 doi: 10.1093/nar/gkab626。 PMID: 34329471; PMCID:PMC8421228。 (影響因子:16.9)
[8] Dai L, Dai Y, Han J, Huang Y, Wang L, Huang J, Zhou Z. BRCA1-BARD1 複合物募集至受損染色質的結構洞察。 Mol Cell。 2021年7月1日;81(13):2765-2777.e6。 doi: 10.1016/j.molcel.2021.05.010。 Epub 2021 年 6 月 7 日。
[9] Zhang K, Wang A, Zhong K, Qi S, Wei C, Shu X, Tu WY, Xu W, Xia C, Xiao Y, Chen A, Bai L, Zhang J, Luo B, Wang W, Shen C. UBQLN2-HSP70 軸減少聚-Gly-AlaOR 聚集體並減輕動物模型中的行為缺陷。神經元。 2021年6月16日;109(12):1949-1962.e6。 doi: 10.1016/j.neuron.2021.04.023。 Epub 2021 年 5 月 14 日。
[10] Liang Y, Lu Q, Li W, Zhang D, Zhang F, Zou Q, Chen L, Tong Y, Liu M, Wang S, Li W, Ren X, Xu P, Yang Z, Dong S, Zhang B, Huang Y, Li D, Wang H, Yu W. 透過 NamiRNA 網路增強子轉換重新活化乳癌中的腫瘤基因轉換。核酸研究。 2021年9月7日;49(15):8556-8572。 doi: 10.1093/nar/gkab626。 PMID: 34329471; PMCID:PMC8421228。 (影響因子:16.9)
[11] Li T, Chen X, Qian Y, Shao J, Li X, Liu S, Zhu L, Zhao Y, Ye H, Yang Y. 一種基於合成BRET的光遺傳學裝置,用於脈衝轉基因表達,實現小鼠葡萄糖穩態。國家通訊社2021年1月27日;12(1):615。 doi: 10.1038/s41467-021-20913-1。 PMID:33504786; PMCID:PMC7840992。 (影響因子:14.92)
[12] Pan Y, He X, Li C, Li Y, Li W, Zhang H, Wang Y, Zhou G, Yang J, Li J, Qu J, Wang H, Gao Z, Shen Y, Li T, Hu H, Ma H. 神經元活動募集 CRTC1/CREB 軸來驅動轉錄依賴性自噬以維持晚期 LTD。 Cell Rep. 2021 年 7 月 20 日;36(3):109398。 doi: 10.1016/j.celrep.2021.109398。 PMID: 34289350. (影響因子:9.42)
[13] Liu H, Xing R, Ou Z, Zhao J, Hong G, Zhao TJ, Han Y, Chen Y. G 蛋白偶聯受體 GPR17 透過增加多梳抑制複合物 1 介導的 ROS 產生來抑制神經膠質瘤發展。細胞死亡論。 2021年6月12日;12(6):610。 doi: 10.1038/s41419-021-03897-0。 PMID: 34120140; PMCID:PMC8197764。 (影響因子:8.463)
[14] 範穎,王建,金偉,孫穎,徐穎,王艷,梁曉燕,蘇丹。分子癌症。 2021年2月2日;20(1):25。 doi: 10.1186/s12943-021-01321-x。PMID: 33530981; PMCID:PMC7851937。 (影響因子:27.403)
[15] 顧 C、王 Y、張 L、喬 L、孫 S、邵 M、唐 X、丁 P、唐 C、曹 Y、週 Y、郭 M、魏 R、李 N、肖 Y、段 J、楊 Y。 J Exp Clin Cancer Res. 2022年1月6日;41(1):11。 doi: 10.1186/s13046-021-02220-1。 PMID: 34991674; PMCID:PMC8734095。 (影響因子:11.161)
[16] 羅倩,吳曉,趙平,南燕,常偉,朱曉,蘇丹,劉哲。 Adv Sci(Weinh)。 2021年2月8日;8(8):2002874。 doi: 10.1002/advs.202002874。 PMID: 33898171; PMCID:PMC8061361。 (影響因子:15.84)
[17] Luo Q, Wu X, Nan Y, Chang W, Zhao P, Zhang Y, Su D, Liu Z.TRIM32/USP11 平衡 ARID1A 穩定性與鱗狀細胞癌的致癌/抑癌狀態。 Cell Rep. 2020年1月7日;30(1):98-111.e5。 doi: 10.1016/j.celrep.2019.12.017。 PMID: 31914402. (影響因子:9.42)
[18] 孫曉玲,彭曉玲,曹穎,週穎,孫穎。國家通訊社2020年6月12日;11(1):2984。 doi: 10.1038/s41467-020-16799-0。 PMID: 32533114; PMCID:PMC7293280。 (影響因子:14.911)
[19] 楊曉玲,王華,謝娥,唐斌,穆倩,宋哲,陳菁,王鋒,閔建軍。蛋白質細胞。 2020年12月;11(12):915-920。 doi: 10.1007/s13238-020-00749-z。 PMID:32632529; PMCID:PMC7719122。 (影響因子:14.872)
[20] 陳 T., 陳 Y., 陳 H. 等.雙酶驅動的無約束DNA步行奈米機器,用於低表達microRNA的細胞內成像。奈米研究12、1055–1060(2019年)。 https://doi.org/10.1007/s12274-019-2344-5 (影響因子:8.21)
[21] 張鑫,齊哲,尹紅,楊剛。治療診斷學。 2019 年 1 月 30 日;9(4):1096-1114。 doi: 10.7150/thno.29673。 PMID: 30867818; PMCID:PMC6401400。 (影響因子:8.12)
[22] Zou Y, Wang A, Shi M, Chen X, Liu R, Li T, Zhang C, Zhang Z, Zhu L, Ju Z, Loscalzo J, Yang Y, Zhao Y. 使用遺傳編碼螢光感測器分析活細胞和體內的氧化還原景觀和動力學。天然協議。 2018年10月;13(10):2362-2386。 doi: 10.1038/s41596-018-0042-5。 PMID: 30258175; PMCID:PMC6714056。 (影響因子:13.49)
[23] 張凱、趙曉、陳曉、魏燕、杜偉、王燕、劉玲、趙偉、韓哲、孔丹、趙倩、郭哲、韓哲、劉娜、馬鋒、李哲。 ACS 應用材料接口。 2018年9月12日;10(36):30081-30091。 doi: 10.1021/acsami.8b08449。 Epub 2018 年 8 月 29 日。
[24]郝海、胡勝、陳華、卜德、朱玲、徐昌、楚鋒、霍曉、唐勇、孫曉、丁本生、劉DP、胡勝、王敏。循環。 2017 年 3 月 28 日;135(13):1253-1264。 doi: 10.1161/CIRCULATIONAHA.116.023027。 Epub 2017 年 2 月 2 日。
付款和安全
您的付款信息已安全處理。我們無法存儲信用卡詳細信息,也無法訪問您的信用卡信息。
詢問
你也可能喜歡
常問問題
本產品僅用於研究目的,不用於人類或動物的治療或診斷用途。產品和內容受以下公司的專利、商標和版權保護:
某些應用程式可能需要額外的第三方智慧財產權。