







설명
Hieff Unicon™ Universal Blue qPCR Master Mix(염료 기반)는 높은 감도와 특이성을 특징으로 하는 2×실시간 정량 PCR 증폭을 위한 사전 솔루션으로, 색상은 파란색이며 샘플 추가 추적 효과가 있습니다. 핵심 구성 요소인 Hieff Unicon™ Taq DNA polymerase는 항체 핫 스타트를 사용하여 샘플 준비 중 프라이머 어닐링으로 인한 비특이적 증폭을 효과적으로 억제합니다. 동시에 이 공식은 PCR 반응의 증폭 효율을 개선하고 다양한 GC 함량(30~70%)을 가진 유전자의 증폭을 균등화하기 위한 촉진 요소를 추가하여 정량 PCR이 광범위한 정량 영역에서 양호한 선형 관계를 얻을 수 있도록 합니다. 이 제품에는 대부분의 qPCR 기기에 적용할 수 있는 특수 ROX Passive Reference Dye가 포함되어 있습니다. 다른 기기에서 ROX 농도를 조정할 필요가 없습니다. 프라이머와 템플릿만 추가하여 증폭을 위한 반응 시스템을 준비하면 됩니다.
특징
- 내장된 시각적 표시기: 실시간 PCR을 방해하지 않고 튜브나 웰에서 더 높은 가시성을 제공하는 불활성 파란색 염료가 포함되어 있습니다.
- 높은 특이성: Taq DNA 중합효소를 포함하고 항체를 통한 열 활성화 메커니즘으로 엄격하게 제어되어 비특이적 증폭을 방지합니다.
- 광범위한 기기 호환성: 대부분의 qPCR 기기와 호환되며 다양한 기기에서 ROX 농도를 조정할 필요가 없습니다.
응용 프로그램
- 유전자 발현
명세서
| 집중 | 2× |
| 검출방법 | SYBR 그린 염료 |
| PCR 방법 | 정자수소핵산분해효소(PCR) |
| 중합효소 | Taq DNA 중합효소 |
| 샘플의 종류 | DNA |
| 응용장비 | 대부분의 qPCR 기기 |
| 제품 유형 | 실시간 형광 정량 PCR을 위한 프리믹스 |
| (신청)에 적용하다 | 유전자 발현 |
구성 요소
| 구성품 번호 | 이름 | 11184ES03 | 11184ES08 | 11184ES50 | 11184ES60 |
| 11184 |
Hieff Unicon™ Universal Blue qPCR 마스터 믹스(염료 기반) |
1ml(1ml) | 5×1ml | 50×1ml | 100×1ml |
배송 및 보관
제품은 아이스팩과 함께 배송되며 -15℃ ~ -25℃에서 18개월 동안 보관할 수 있습니다. 제품에는 형광 염료가 포함되어 있으므로 반응 시스템을 보관하거나 제조할 때 강한 빛 조사를 피해야 합니다.
그림
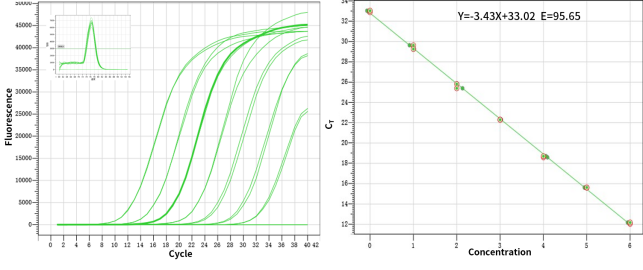
그림 1. 높은 민감도: 단일 사본을 감지하는 능력.
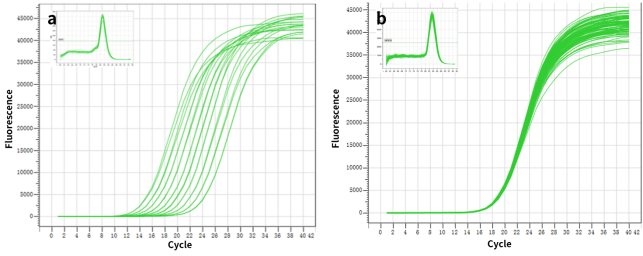
그림 2. 복제 웰의 높은 분해능 및 우수한 재현성.
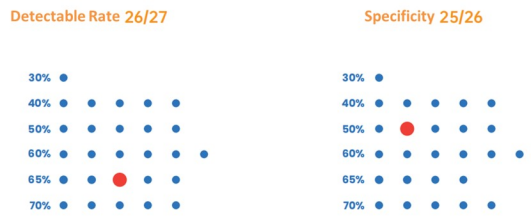
그림 3. 우수한 검출률과 특이성: 30-70% GC 증폭에 광범위하게 적합함.

"SARS-CoV-2 봉투 단백질은 급성 호흡곤란증후군(ARDS)과 유사한 병리적 손상을 일으키고 항바이러스 표적을 구성합니다. Cell Res. 2021;31(8):847-860. doi:10.1038/s41422-021-00519-4"에서 인용

"플라스마 막과 엽록체를 연결하고 병원균에 의해 채택된 방어 경로. Cell. 2020년 9월 3일;182(5):1109-1124.e25. doi: 10.1016/j.cell.2020.07.020"에서 인용.
[1] Xia B, Shen X, He Y, et al. SARS-CoV-2 봉투 단백질은 급성 호흡곤란증후군(ARDS)과 유사한 병리학적 손상을 유발하고 항바이러스 표적을 구성합니다. Cell Res. 2021;31(8):847-860. doi:10.1038/s41422-021-00519-4(IF:25.617)
[2] Wang S, Li Y, Zhong L, et al. 세포 내 인트론 선택 마커를 통한 효율적인 유전자 편집. Cell Mol Life Sci. 2022;79(2):111. 2022년 1월 31일 출판. doi:10.1007/s00018-022-04152-1(IF:9.261)
[3] An LL, Zhao X, Gong XY, et al. IFN 반응에서 Zebrafish IRF Family Members의 프로모터 결합 및 핵 유지 특징. Front Immunol. 2022;13:861262. 2022년 4월 6일 출판. doi:10.3389/fimmu.2022.861262(IF:7.561)
[4] Zhao Y, Wang HP, Yu C, et al. 유도적 스트레스 조건에서 Haematococcus pluvialis의 아스타잔틴 및 지질 공동 생산에 대한 멜라토닌의 자극 효과의 근저에 있는 조절 메커니즘을 밝히기 위한 생리학적 및 대사체학적 프로필 통합. Bioresour Technol. 2021;319:124150. doi:10.1016/j.biortech.2020.124150(IF:7.539)
[5] Shu C, Wang L, Zou C, et al. 올리브 넙치 Paralichthys olivaceus의 생식소 분화 중 Foxl2 및 Dmrt1 단백질의 기능 [2022년 6월 16일 인쇄 전 온라인 출판]. Int J Biol Macromol. 2022;215:141-154. doi:10.1016/j.ijbiomac.2022.06.098(IF:6.953)
[6] Wang S, Huang J, Liu F, et al. Isosteviol Sodium Exerts Anti-Colitic Effects on BALB/c Mice with Dextran Sodium Sulfate-Induced Colitis Through Metabolic Reprogramming and Immune Response Modulation. J Inflamm Res. 2021;14:7107-7130. 2021년 12월 20일 출판. doi:10.2147/JIR.S344990(IF:6.922)
[7] Wang J, Hu R, Wang Z, et al. 렌티바이러스 매개 SV40T 및 hTERT 유전자 전달에 의한 불멸화된 야크 반추위 상피 세포주 확립. Oxid Med Cell Longev. 2022;2022:8128028. 2022년 3월 25일 출판. doi:10.1155/2022/8128028(IF:6.543)
[8] Liu W, Guan Y, Qiao S, et al. Vicatia thibetica de Boiss 뿌리 추출물의 Caenorhabditis elegans 및 성인 마우스의 독소루비신 유도 조기 노화에 대한 노화 방지 효과. Oxid Med Cell Longev. 2021;2021:9942090. 2021년 8월 6일 출판. doi:10.1155/2021/9942090(IF:6.543)
[9] Qian Z, Liu C, Li H, et al. Osteocalcin Alleviates Lipopolysaccharide-Induced Acute Inflammation via Activation of GPR37 in Macrophages. Biomedicines. 2022;10(5):1006. 2022년 4월 27일 출판. doi:10.3390/biomedicines10051006(IF:6.081)
[10] Zhao X, Huang Y, Li X, et al. 키토산으로 변형된 미세구체를 사용한 원심 마이크로유체 칩으로의 핵산 추출 및 검출의 완전 통합 [2022년 6월 27일 인쇄 전 온라인 출판]. Talanta. 2022;250:123711. doi:10.1016/j.talanta.2022.123711(IF:6.057)
[11] Xu X, Wang H, Li X, Duan X, Wang Y. 새로운 ALG10/TGF-β 양성 조절 루프는 대장암의 줄기세포성에 기여합니다. 노화(Albany NY). 2022;14(11):4858-4873. doi:10.18632/aging.204116(IF:5.955)
[12] Li F, Hu X, Wu J. Daidzein은 Clec11a를 상향 조절하여 여성 생식세포 줄기세포의 증식을 촉진하기 위해 Akt 경로를 활성화합니다[2022년 6월 3일 인쇄 전 온라인 출판]. Stem Cell Rev Rep. 2022;10.1007/s12015-022-10394-0. doi:10.1007/s12015-022-10394-0(IF:5.739)
[13] Ru M, Wang W, Zhai Z, et al. 니코틴아마이드 모노뉴클레오타이드 보충은 노화 마우스와 D-갈락토스 유도 노화 세포에서 장 기능을 보호합니다. 식품 기능. 2022;13(14):7507-7519. 2022년 7월 18일 출판. doi:10.1039/d2fo00525e(IF:5.396)
[14] Ye M, Gao R, Chen S, et al. MEG3의 다운레귤레이션과 EZH2의 업레귤레이션은 협력적으로 신경아세포종 진행을 촉진합니다. J Cell Mol Med. 2022;26(8):2377-2391. doi:10.1111/jcmm.17258(IF:5.310)
[15] Wang S, Wang X, Shao Y, et al. 새로운 HIF-1 억제제로서 3-(페닐에티닐)-1,1'-비페닐-2-카복실레이트 유도체의 합성 및 평가. Bioorg Chem. 2021;116:105298. doi:10.1016/j.bioorg.2021.105298(IF:5.275)
[16] Guan X, Meng X, Zhu K, et al. MYSM1은 RSK3-phospho-BAD 경로를 통해 세포사멸을 유도하고 TNBC 세포를 시스플라틴에 민감하게 만듭니다. Cell Death Discov. 2022;8(1):84. 2022년 2월 26일 출판. doi:10.1038/s41420-022-00881-1(IF:5.241)
[17] Liu H, Wang LL, Xu QH, et al. UHRF1은 임신 초기에 태반모세포 침윤과 낙엽세포 대식세포 분화를 모두 형성합니다. FASEB J. 2022;36(4):e22247. doi:10.1096/fj.202101647RR(IF:5.192)
[18] Chen M, Wang J, Lin L, et al. Penicillium brevicompactum의 Ca<sup>2+</sup>/반응성 산소 종에 의한 대사의 상승적 조절은 미코페놀산 생성을 개선하고 Ca<sup>2+</sup> 채널에 대한 연구를 개선합니다 [게시된 수정 사항은 ACS Synth Biol. 2022년 4월 15일;11(4):1705에 나타남]. ACS Synth Biol. 2022년;11(1):273-285. doi:10.1021/acssynbio.1c00413(IF:5.110)
[19] Zhao X, Li X, Yang W, Peng J, Huang J, Mi S. 고위험 인유두종바이러스의 자동화 및 신속한 진단을 위한 통합 마이크로유체 감지 시스템. Analyst. 2021;146(16):5102-5114. doi:10.1039/d1an00623a(IF:4.616)
[20] Chen H, Jiang Y, Liu R, et al. Curcumin Derivative C66 Suppresses Pancreatic Cancer Progression through the Inhibition of JNK-Mediated Inflammation. Molecules. 2022;27(10):3076. 2022년 5월 11일 출판. doi:10.3390/molecules27103076(IF:4.412)
[21] Su W, Qiu J, Mei Y, Zhang XE, He Y, Li F. 허용 세포에 대한 신속한 스크리닝을 통한 바이러스 분리를 위한 마이크로유체 세포 칩 [2022년 5월 2일 인쇄 전 온라인 출판]. Virol Sin. 2022;S1995-820X(22)00075-X. doi:10.1016/j.virs.2022.04.011(IF:4.327)
[22] Ding J, Mei S, Cheng W, Ni Z, Yu C. 커큐민은 HIF 신호 전달 경로를 통해 생쥐의 자궁 내막증을 치료합니다. Am J Transl Res. 2022;14(4):2184-2198. 2022년 4월 15일 출판. (IF:4.060)
[23] Zhang XY, Shen HH, Qin XY, et al. IL-27은 STAT3-ESR/PGR 조절 축을 통해 탈락막화를 촉진합니다. J Reprod Immunol. 2022;151:103623. doi:10.1016/j.jri.2022.103623(IF:4.054)
[24] Zhu Q, Muyayalo KP, Xu QH, Wang J, Wang H, Liao AH. Prunella vulgaris는 Th1/Th17 면역 반응을 억제하여 실험적 자가면역성 갑상선염 쥐의 임신 결과를 개선할 수 있습니다. J Reprod Immunol. 2022;149:103469. doi:10.1016/j.jri.2021.103469(IF:4.054)
[25] Yang Q, Guo K, Zhou X, et al. 세균 감염 후 삼각돛 조개 Hyriopsis cumingii의 조직병리학, 항산화 반응, 전사체 및 유전자 발현 분석. Dev Comp Immunol. 2021;124:104175. doi:10.1016/j.dci.2021.104175(IF:3.636)
[26] Liu Y, Wan HH, Tian DM, et al. 유전자 치료를 위한 히스티딘 및 아르기닌 비율 조절을 통한 고효율 세포 침투 펩타이드의 개발 및 특성화. Materials(Basel). 2021;14(16):4674. 2021년 8월 19일 출판. doi:10.3390/ma14164674(IF:3.623)
[27] Fu Y, Wang X, Zhang L, Ren Y, Hao L. 이식편 염증 인자-1은 당뇨병 신장 질환에서 NF-κB 경로를 통해 염증과 산화 스트레스를 증가시킵니다. Biochem Biophys Res Commun. 2022;614:63-69. doi:10.1016/j.bbrc.2022.04.089(IF:3.575)
[28] Zhao Y, Wang XQ. VvMYB1은 VvTOR 프로모터를 조절하여 VvTOR 유전자 발현에 영향을 미칠 가능성이 있으며 포도당 축적에 참여합니다. J Plant Physiol. 2022;272:153668. doi:10.1016/j.jplph.2022.153668(IF:3.549)
[29] Cheng Y, Zheng L, Yang C, Zhang W, Wang H. Propofol은 lncRNA GAS5를 상향 조절하여 신경교종 세포의 증식과 이동을 억제합니다. Toxicol In Vitro. 2022;80:105321. doi:10.1016/j.tiv.2022.105321(IF:3.500)
[30] Yang B, Wang F, Zheng G. 막관통 단백질 TMEM119는 Wnt/β-catenin 경로를 활성화하여 유방암 세포의 줄기세포성을 촉진합니다. Bioengineered. 2021;12(1):4856-4867. doi:10.1080/21655979.2021.1960464(IF:3.269)
[31] Yang X, Liu X, Song F, et al. 야생 땅 다람쥐(<em>Spermophilus dauricus</em>) 대장에서의 GPR41 및 GPR43의 계절적 발현. Eur J Histochem. 2022;66(1):3351. 2022년 1월 21일 출판. doi:10.4081/ejh.2022.3351(IF:3.188)
[32] Tang Z, Yuan X, Bai Y, et al. 사향쥐(<em>Ondatra zibethicus</em>) 고환의 PACAP, VPAC1, VPAC2, PAC1 발현 및 고환 활동의 계절적 변화. Eur J Histochem. 2022;66(2):3398. 2022년 5월 2일 출판. doi:10.4081/ejh.2022.3398(IF:3.188)
[33] Zhu P, Chen Y, Wu F, Meng M, Ji K. Pinus massoniana Lamb에서 MEP 경로 효소 인코딩 유전자의 발현 및 프로모터 분석. PeerJ. 2022;10:e13266. 2022년 4월 12일 출판. doi:10.7717/peerj.13266(IF:2.984)
[34] Zhai L, Chen W, Cui B, Yu B, Wang Y, Liu H. 과잉 발현된 베르시칸은 위암에서 세포 증식, 이동 및 침윤을 촉진합니다. 조직 세포. 2021;73:101611. doi:10.1016/j.tice.2021.101611(IF:2.466)
[35] Jiang J, Cai M. Cardamonin은 연골세포에서 Nrf2/NQO-1 신호 전달 경로를 활성화하여 NLRP3 인플라마좀을 억제하여 IL-1β 유도 손상을 억제했습니다. J Microbiol Biotechnol. 2021;31(6):794-802. doi:10.4014/jmb.2103.03057(IF:2.351)
결제 및 보안
귀하의 결제 정보는 안전하게 처리됩니다. 당사는 신용카드 정보를 저장하지 않으며 귀하의 신용카드 정보에 접근할 수 없습니다.
문의
당신은 또한 좋아할 수 있습니다
자주 묻는 질문
이 제품은 연구 목적으로만 사용되며 인간이나 동물의 치료 또는 진단용으로 의도되지 않았습니다. 제품과 콘텐츠는
특정 애플리케이션에는 추가적인 제3자 지적 재산권이 필요할 수 있습니다.
예슨은 윤리적 과학에 헌신하며, 우리의 연구가 안전과 윤리적 기준을 보장하는 동시에 중요한 문제를 해결해야 한다고 믿습니다.