

설명
GMyc-PCR 마이코플라스마 검출 키트는 주로 PCR 방법을 사용하여 다양한 생물학적 물질(예: 세포 배양, 실험 동물 분비물, 동물 혈청 등)의 마이코플라스마 감염을 검출합니다. 민감하고, 특이적이며, 빠르며, 세포 배양 상층액으로 직접 검출할 수 있는 여러 가지 장점이 결합되어 있습니다. 이 제품은 PCR 방법으로 배양 세포와 같은 생물학적 물질에서 마이코플라스마를 검출합니다. 사용된 프라이머는 마이코플라스마의 16S-23S rRNA 서열의 보존 영역에 따라 설계되었으며, 마이코플라스마 DNA만 특이적으로 증폭하여 검출 감도와 특이성이 높습니다. PCR 증폭 및 전기영동 분석은 몇 시간 밖에 걸리지 않으며 조작이 편리하고 간단합니다.
세포 배양은 생명 과학 연구에서 일반적인 실험입니다. 일반적으로 사용되는 다른 실험 방법과 달리 세포 배양은 역동적인 연속적 과정이며 세포는 종종 비정상적인 세포 상태 또는 배지 모양을 나타내는 조작 오류 또는 오염 물질에 반응합니다. 마이코플라스마에 오염된 경우 세포 형태에는 눈에 띄는 변화가 없으며 간과하기 쉽습니다. 오염이 매우 심각해질 때까지 종종 발견되지 않습니다. 오염된 세포막에는 수백 개의 마이코플라스마가 있을 수 있으며 이러한 마이코플라스마는 영양소를 놓고 경쟁하고 독성 대사 산물을 방출하여 실험 결과에 심각한 영향을 미칩니다.
연구에 따르면 최소 20종의 마이코플라스마가 세포를 오염시킬 수 있으며, 그 중 가장 흔한 것은 다음과 같습니다.경구 마이코플라스마(M. orale), 마이코플라스마 아르기닌( M. arginini ), 마이코플라스마 하이오리니스( M. hyorhinis ) , 마이코플라스마 페르멘툼( M. fermentans ), 마이코플라스마 호미니스( M. hominis ), 마이코플라스마 살리바리우스( M. salivarium), 마이코플라스마 폐(M. pulmonis ), 마이코플라스마 배( M. pirum ).배양된 세포의 마이코플라스마 오염률은 4%에서 92% 사이입니다.오염원에는 작업 환경, 작업자 본인(일부 마이코플라스마는 인체의 정상 세균총), 배양 배지, 혈청, 세포 교차 오염, 실험 장비, 세포가 준비된 원래 조직이나 장기의 오염이 포함됩니다.
세포 배양 중 문제의 근본 원인을 파악하는 것은 어렵고 시간이 많이 걸리는 작업으로, 갑작스러운 변화가 의심되어야 하며, 마이코플라스마 오염에 대한 우수한 검사 관행과 정기적인 검사가 필요합니다. 마이코플라스마를 검출하는 방법에는 직접 배양, DNA 형광 염색, ELISA, PCR 방법 등 여러 가지가 있습니다.
예센 마이코플라스마 오염에 대한 포괄적인 솔루션을 제공합니다 . 관련 제품을 살펴보세요: 마이코플라스마 개념과 오염의 영향
특징
- 사용된 프라이머는 Mycoplasma 16S-23S rRNA 서열의 보존 영역에 따라 설계되었습니다.
- 증폭된 마이코플라스마 DNA만을 특별히
- 뛰어난 민감도와 특이성
- 마이코플라스마 종 20종 이상이 검출됐다.
애플리케이션
- 마이코플라스마 검출
구성 요소
| 구성품 번호 | 이름 | 40601ES10(10개 분석) | 40601ES20(20개 분석) |
| 40601-A | GMyc-1st PCR 믹스 | 250µL | 2×250µL |
| 40601-B | GMyc-2nd PCR 믹스 | 250µL | 2×250µL |
| 40601-C | 양성 대조 템플릿 N | 20µL | 20µL |
[주의사항] 1. 장기간 사용하지 않을 경우 -85~-65℃에서 냉동보관 가능합니다.
2. PCR 반응은 매우 민감합니다. 거짓 양성을 방지하기 위해 샘플을 추가할 때 끝에 양성 대조군을 추가합니다.
저장
본 제품은 -25~-15℃에서 18개월간 보관 가능합니다. 장기간 사용하지 않을 경우 빛으로부터 멀리 보관해 주세요.
그림
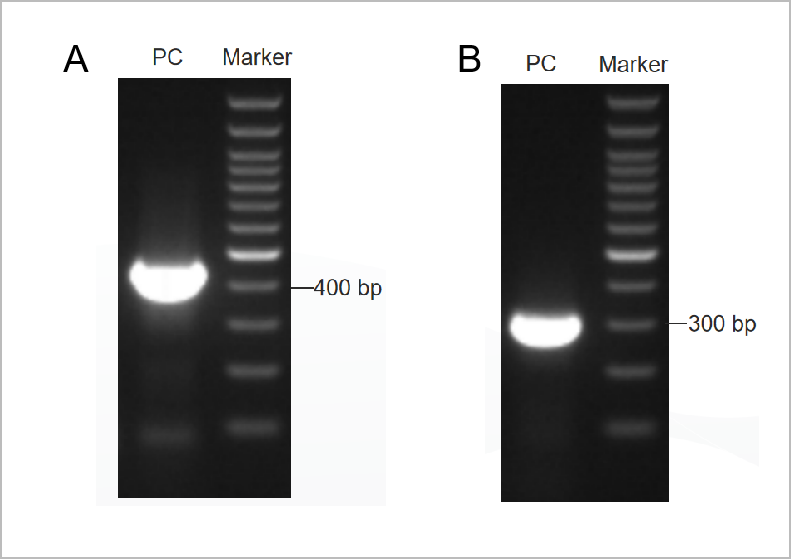
그림 1. PCR Mycoplasma Test Kit의 전기영동 결과 .
첫 번째 라운드 전기영동도는 Figure 1A에 표시되어 있으며 두 번째 라운드 전기영동도는 Figure 1B에 표시되어 있습니다.(M: 1kb 마커, 1-2: 양성 대조군, 3: 음성 대조군)
[1] Rao XS, Cong XX, Gao XK, et al. AMPK 매개 인산화는 Rab5 의존적 포도당 흡수를 촉진하기 위해 TBC1D17의 자동 억제를 향상시킵니다. Cell Death Differ. 2021;28(12):3214-3234. doi:10.1038/s41418-021-00809-9(IF:15.828)
[2] Guo F, Li L, Li J, et al. 마우스 초기 배아 및 배아줄기세포의 단일세포 다중오믹스 시퀀싱. Cell Res. 2017;27(8):967-988. doi:10.1038/cr.2017.82(IF:15.606)
[3] Hao Y, He B, Wu L, et al. p85β의 핵 전좌는 PIK3CA 나선형 도메인 돌연변이 암의 종양 형성을 촉진합니다. Nat Commun. 2022;13(1):1974. 2022년 4월 13일 출판. doi:10.1038/s41467-022-29585-x(IF:14.919)
[4] Shu X, Liu M, Lu Z, et al. 게놈 전체 매핑을 통해 데옥시우리딘이 인간 중심체 DNA에 풍부하다는 사실이 밝혀졌습니다. Nat Chem Biol. 2018;14(7):680-687. doi:10.1038/s41589-018-0065-9(IF:13.843)
[5] Li X, Xiong X, Wang K, et al. 전사체 전체 매핑은 가역적이고 동적인 N(1)-메틸아데노신 메틸롬을 밝힙니다. Nat Chem Biol. 2016;12(5):311-316. doi:10.1038/nchembio.2040(IF:12.709)
[6] Sun L, Yang X, Huang X, et al. 지방산의 2-하이드록실화는 YAP 전사 축을 통해 대장암 종양 형성 및 전이를 억제합니다. Cancer Res. 2021;81(2):289-302. doi:10.1158/0008-5472.CAN-20-1517(IF:12.701)
[7] Sun Z, Zhang Z, Wang QQ, Liu JL. CTPS1과 ATR의 결합 불활성화는 MYC 과발현 암세포에 합성적으로 치명적입니다. Cancer Res. 2022;82(6):1013-1024. doi:10.1158/0008-5472.CAN-21-1707(IF:12.701)
[8] Song J, Zhuang Y, Zhu C, et al. miRNA 처리 및 tRNA 의사유리딜화에서 인간 PUS10의 차별적 역할. Nat Chem Biol. 2020;16(2):160-169. doi:10.1038/s41589-019-0420-5(IF:12.154)
[9] He B, Pan H, Zheng F, et al. 긴 비번역 RNA LINC00930은 비인두암에서 PFKFB3 매개 종양 해당분해 및 세포 증식을 촉진합니다. J Exp Clin Cancer Res. 2022;41(1):77. 2022년 2월 24일 출판. doi:10.1186/s13046-022-02282-9(IF:11.161)
[10] Tang B, Liu BH, Liu ZY, Luo MY, Shi XH, Pang DW. 컴팩트한 친수성 쌍성 이온 코팅을 갖춘 양자점. ACS Appl Mater Interfaces. 2022;14(24):28097-28104. doi:10.1021/acsami.2c04438(IF:9.229)
[11] Huang C, Zhang Z, Chen L, et al. Src의 N 및 C 말단 도메인 내 아세틸화는 STAT3 매개 종양 형성의 고유한 역할을 조절합니다. Cancer Res. 2018;78(11):2825-2838. doi:10.1158/0008-5472.CAN-17-2314(IF:9.130)
[12] Wu X, Yu M, Zhang Z, et al. DDB2는 CDT2의 PCNA 독립적 분해를 통해 DNA 복제를 조절합니다. Cell Biosci. 2021;11(1):34. 2021년 2월 8일 출판. doi:10.1186/s13578-021-00540-5(IF:7.133)
[13] Wang J, Zhang Y, Liu X, Liu H. 종양 저항성 부분 집단에 대한 도달 가능성에 기반한 적응 치료 최적화. Cancers(Basel). 2021;13(21):5262. 2021년 10월 20일 출판. doi:10.3390/cancers13215262(IF:6.639)
[14] Feng W, Liu R, Xie X, et al. α-tubulin의 SUMOylation은 미세소관 역학을 조절하는 새로운 변형입니다. J Mol Cell Biol. 2021;13(2):91-103. doi:10.1093/jmcb/mjaa076(IF:6.216)
[15] Yu M, Hu X, Yan J, Wang Y, Lu F, Chang J. RIOK2 억제제 NSC139021은 Skp2 매개 세포주기 정지 및 세포사멸 유도를 통해 신경교종에 항종양 효과를 발휘합니다. Biomedicines. 2021;9(9):1244. 2021년 9월 17일 출판. doi:10.3390/biomedicines9091244(IF:6.081)
[16] Ren S, Cai Y, Hu S, et al. Berberine은 c-myc/CD47 축을 조절하여 확산성 대형 B세포 림프종에서 항종양 활동을 발휘합니다. Biochem Pharmacol. 2021;188:114576. doi:10.1016/j.bcp.2021.114576(IF:5.858)
[17] Wen F, Sun X, Sun C, et al. TAGLN은 전립선암 세포에서 TRAF6 매개 프로테아좀 분해에 의해 다운레귤레이션됩니다. Mol Cancer Res. 2021;19(7):1113-1122. doi:10.1158/1541-7786.MCR-20-0513(IF:5.852)
[18] Tang B, Sun EZ, Zhang ZL, et al. 스핑고미엘린 격리 콜레스테롤 도메인은 인플루엔자 바이러스 진입 시 클라트린 코팅 구덩이를 수축시키기 위해 포르민 결합 단백질 17을 모집합니다. J Virol. 2022;96(5):e0181321. doi:10.1128/JVI.01813-21(IF:5.103)
[19] Hu J, Ren W, Qiu W, et al. JAK2 돌연변이가 있는 강직성 척추염 환자로부터 유도 다능성 줄기 세포주(XDCMHi001-A) 생성. Stem Cell Res. 2020;45:101788. doi:10.1016/j.scr.2020.101788(IF:4.495)
[20] Xiao S, Yao X, Ye J, Tian X, Yin Z, Zhou L. 후성유전적 변형은 위암에서 프로린 합성효소 PYCR1의 이상 발현을 촉진합니다 [2022년 5월 30일 인쇄 전 온라인 출판]. Biochim Biophys Acta Gene Regul Mech. 2022;1865(6):194829. doi:10.1016/j.bbagrm.2022.194829(IF:4.490)
[21] Wang J, Zhang Y, Liu X, Liu H. 완전한 정보 없이 고정 주기 치료가 종양 시스템에 효과적일까요?. Cancer Manag Res. 2021;13:8915-8928. 2021년 11월 30일 출판. doi:10.2147/CMAR.S339787(IF:3.989)
[22] Yang X, Ren S, Rehman ZU, et al. 소(Bos taurus) 및 염소(Capra hircus)의 TANK-결합 키나제 1(TBK1)의 분자적 특성화, 발현 및 기능 식별. Dev Comp Immunol. 2022;133:104444. doi:10.1016/j.dci.2022.104444(IF:3.636)
[23] Zheng D, Chang X, Liu Y, et al. 2-Methoxy-5((3,4,5-trimethosyphenyl)seleninyl) phenol은 유방암 세포주에서 MDM2의 다운레귤레이션을 통해 EGF 유도 세포 이동 및 침습을 역전시킵니다. Cancer Biol Ther. 2019;20(4):513-523. doi:10.1080/15384047.2018.1537578(IF:3.373)
[24] Xu F, Zhang S, Liu Z, et al. TEX9와 eIF3b는 기능적으로 상승작용하여 식도 편평세포 암의 진행을 촉진합니다. BMC Cancer. 2019;19(1):875. 2019년 9월 3일 출판. doi:10.1186/s12885-019-6071-9(IF:2.933)
[25] Pan H, Sun L, Wang W, et al. 혈청 긴 비코딩 RNA LOC553103은 일반적인 인간 암 유형에 대한 비특이적 진단 및 예후 바이오마커입니다. Clin Chim Acta. 2020;508:69-76. doi:10.1016/j.cca.2020.05.017(IF:2.615)
[26] Li N, Lin SM, Li Y, Sun J, Zhang L, Chen M. FGFR3 유전자에 이형접합 G380R 돌연변이가 있는 태아로부터 유래한 유도만능줄기세포주(GZHMCi004-A)는 연골무형성을 유발합니다. Stem Cell Res. 2021;53:102322. doi:10.1016/j.scr.2021.102322(IF:2.020)
[27] Luo Q, Wei C, Long Y, et al. iCRISPR/Cas9 시스템을 사용한 ELTD1 녹아웃 인간 배아줄기세포주 생성. Stem Cell Res. 2021;53:102350. doi:10.1016/j.scr.2021.102350(IF:2.020)
[28] Liu YQ, Ling TW, Wang HY, Yang YH, Song WJ, Wang TC. DGKG 돌연변이가 있는 간질 환자로부터 통합 없는 유도 다능성 줄기 세포주(LZUSHI001-A) 생성. Stem Cell Res. 2022;61:102768. doi:10.1016/j.scr.2022.102768(IF:2.020)
[29] Chen M, Lin SM, Li N, Li Y, Li Y, Zhang L. RUNX2 유전자의 엑손 3 이형 결손으로 인해 쇄골두개골 이형성을 유발하는 태아로부터 유래한 유도 다능성 줄기 세포주(GZHMCi003-A). Stem Cell Res. 2021;51:102166. doi:10.1016/j.scr.2021.102166(IF:2.020)
[30] Xu Y, Wang X, Qiu T, et al. DNM1 유전자에 이형접합 c.545C > A 돌연변이를 갖는 발달성 및 간질성 뇌병증 31형 환자로부터 유래한 유도 다능성 줄기 세포주(FDCHI007-A) 생성. Stem Cell Res. 2022;60:102709. doi:10.1016/j.scr.2022.102709(IF:2.020)
[31] Fan T, He J, Wang Y, Yu J, Sun W. 중추성 조숙성 사춘기를 가진 7세 여아로부터 유도 다능성 줄기 세포주(FDCHi006-A) 생성. Stem Cell Res. 2021;56:102542. doi:10.1016/j.scr.2021.102542(IF:2.020)
[32] Gong X, Zheng Z, Yang T, Zheng H, Xiao X, Jia N. ARX 유전자의 반접합 R330L 돌연변이가 있는 1형 조기 유아 간질성 뇌병증(EIEE1) 환자로부터 등유전자 교정 iPSC 계열(OGHFUi001-A-1) 생성. 줄기세포 연구. 2022;60:102693. doi:10.1016/j.scr.2022.102693(IF:2.020)
[33] Jia N, Gong X, Chen J, et al. ARX 돌연변이가 있는 1형 유아 초기 간질성 뇌병증으로부터 유도 다능성 줄기 세포주(OGHFUi001-A) 생성. Stem Cell Res. 2021;53:102367. doi:10.1016/j.scr.2021.102367(IF:2.020)
[34] Zhu W, Zhou Y, Wang Q, et al. PSEN1 유전자에 E363Q 돌연변이를 지닌 환자의 피부 섬유아세포로부터 인간 유도 다능성 줄기 세포(iPSC) 세포주 생성. Stem Cell Res. 2022;61:102769. doi:10.1016/j.scr.2022.102769(IF:2.020)
[35] Luo F, Long K, Li X, et al. LRRC8A 결핍은 3T3-L1 세포에서 저산소 유도 괴사를 약화시킵니다. Biosci Biotechnol Biochem. 2020;84(6):1139-1145. doi:10.1080/09168451.2020.1730689(IF:1.516)
결제 및 보안
귀하의 결제 정보는 안전하게 처리됩니다. 당사는 신용카드 정보를 저장하지 않으며 귀하의 신용카드 정보에 접근할 수 없습니다.
문의
당신은 또한 좋아할 수 있습니다
자주 묻는 질문
이 제품은 연구 목적으로만 사용되며 인간이나 동물의 치료 또는 진단용으로 의도되지 않았습니다. 제품과 콘텐츠는
특정 애플리케이션에는 추가적인 제3자 지적 재산권이 필요할 수 있습니다.
예슨은 윤리적 과학에 헌신하며, 우리의 연구가 안전과 윤리적 기준을 보장하는 동시에 중요한 문제를 해결해야 한다고 믿습니다.