





Sự miêu tả
Hạt chọn lọc DNA Hieff NGS™ được chế tạo dựa trên nguyên lý SPRI (Bất động hóa ngược pha rắn) và được áp dụng để tinh chế DNA và chọn kích thước trong quá trình chuẩn bị thư viện giải trình tự thế hệ tiếp theo (NGS). Hạt chọn lọc DNA Hieff NGS™ tương thích với nhiều bộ dụng cụ chuẩn bị thư viện DNA và RNA và là một giải pháp thay thế tốt Hạt AMPure.
Thành phần
| Số thành phần | Tên | 12601ES08 | 12601ES56 | 12601ES75 |
| 12601 | Hạt lựa chọn DNA Hieff NGS™ | 5ml | 60ml | 450mL |
Thông số kỹ thuật
| Dòng sản phẩm | Hạt DNA sạch và chọn lọc |
| Vật liệu khởi đầu | ADN |
| Khả năng tương thích | ADN |
| Công nghệ cách ly | Hạt từ tính |
| Loại sản phẩm cuối cùng | ADN |
| Để sử dụng với (Ứng dụng) | DNA celan up, DNA size slection |
Vận chuyển và lưu trữ
Các hạt được vận chuyển bằng túi đá và có thể bảo quản ở nhiệt độ 2°C-8°C trong một năm.
Hướng dẫn
- 1. Chuẩn bị
Để các hạt lựa chọn ở nhiệt độ phòng trong ít nhất 30 phút trước khi sử dụng.
- 2. Lựa chọn kích thước DNA
Luồng hoạt động lựa chọn kích thước được thể hiện ở Hình 1 và giao thức như sau.
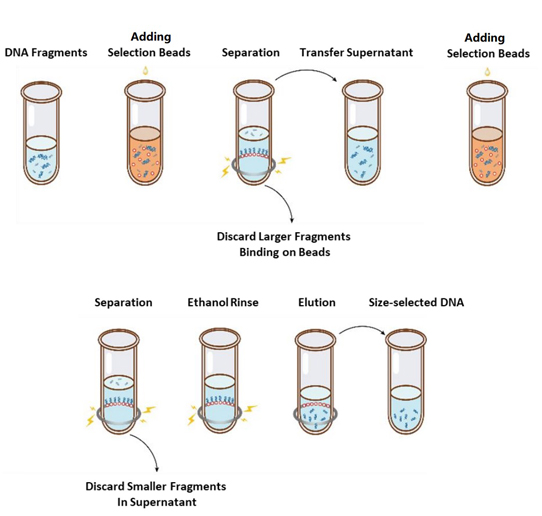
Hình 1. Sơ đồ dòng chảy của việc lựa chọn kích thước DNA
2.1 Trộn đều các hạt bằng cách lắc hoặc hút lên xuống mỗi lần trước khi sử dụng.
2.2 Thêm vòng hạt chọn lọc đầu tiên vào mẫu (tham khảo Bảng 1). Trộn đều bằng cách lắc xoáy hoặc hút lên xuống ít nhất 10 lần.
2.3 Ủ ở nhiệt độ phòng trong 5 phút.
2.4 Xoay ống xuống một chút và đặt lên giá từ. Khi dung dịch trong (khoảng 5 phút), chuyển phần dịch nổi vào ống PCR mới.
2.5 Thêm vòng hạt chọn lọc thứ hai vào mẫu từ bước 2.4 theo Bảng 1. Trộn đều bằng cách lắc hoặc hút lên xuống ít nhất 10 lần.
2.6 Ủ ở nhiệt độ phòng trong 5 phút.
2.7 Xoay ống xuống một lát và đặt lên giá từ. Khi dung dịch trong (khoảng 5 phút), hút phần chất lỏng trong và loại bỏ.
2.8 Giữ ống trong giá từ và thêm 200 μL ethanol 80% mới pha vào mà không làm xáo trộn các hạt, ủ ở nhiệt độ phòng trong 30 giây. Hút ethanol và loại bỏ.
2.9 Lặp lại bước 2.8 một lần để giặt tổng cộng hai lần.
2.10 Loại bỏ etanol còn lại bằng đầu pipet 10 µL. Giữ ống trong giá đỡ từ tính, phơi khô các hạt lựa chọn bằng không khí với nắp mở cho đến khi các vết nứt xuất hiện (khoảng 5 phút).
Lưu ý: Không làm khô các hạt lựa chọn quá mức. Điều này có thể làm giảm mục tiêu phục hồi DNA.
2.11 Lấy ống ra khỏi giá từ. Thêm một lượng ddH2O thích hợp (≥20 µL) và trộn đều bằng cách vortex hoặc hút lên xuống ít nhất 10 lần.
2.12 Ủ ở nhiệt độ phòng trong 5 phút.
Xoay ống xuống một lát và đặt lên giá từ. Khi dung dịch trong (khoảng 5 phút), chuyển 20 μL dịch trong sang ống mới.
- 3. Điều kiện khuyến nghị để lựa chọn kích thước DNA
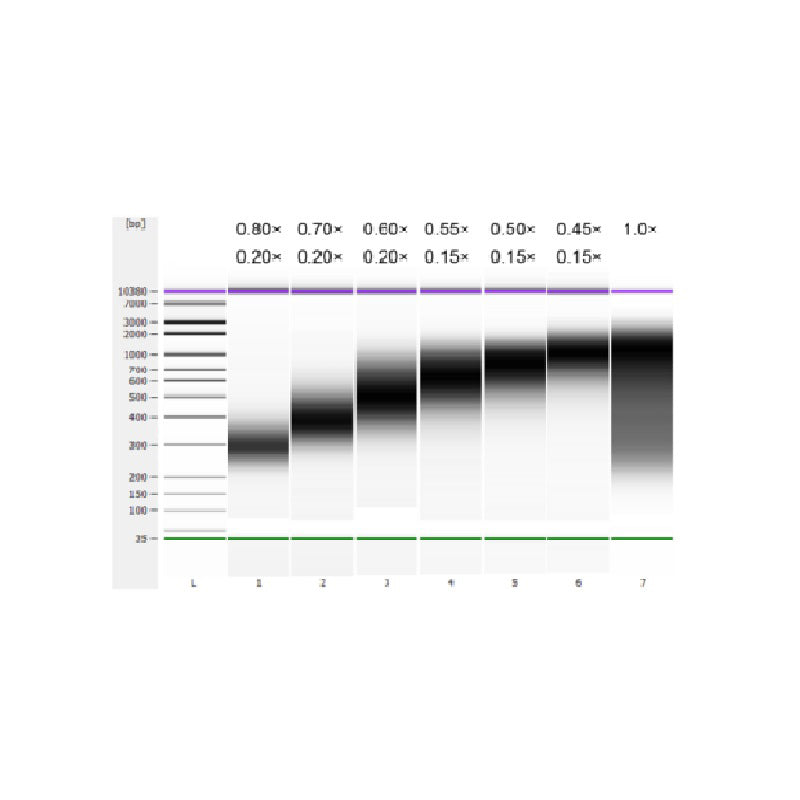
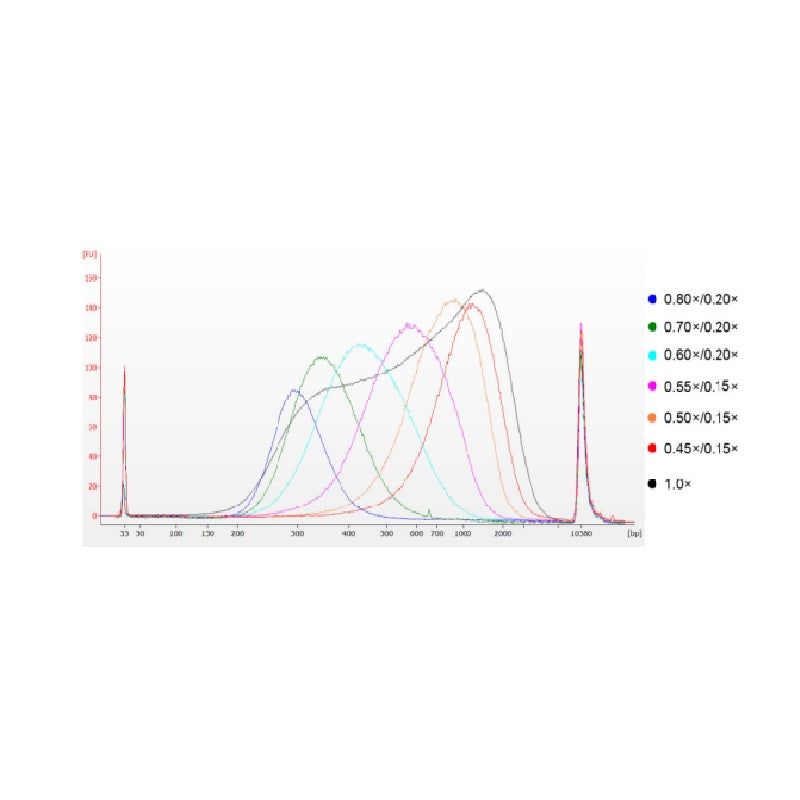
DNA tuyến ức bê được phân mảnh bằng phương pháp siêu âm để chuẩn bị một đoạn có kích thước 100-1.000 bp và thực hiện hai vòng lựa chọn kích thước theo Bảng 1. Kết quả được phân tích bằng Agilent 2100 Bioanalyzer (Hình 2).
Bảng 1. Điều kiện khuyến nghị để lựa chọn kích thước DNA
| Chiều dài của đoạn DNA | 250-350 điểm chuẩn | 320-420 điểm chuẩn | 450-550 điểm chuẩn | 550-700 điểm chuẩn | 700-900 điểm chuẩn | 800-1.000 điểm chuẩn |
| Tỷ lệ hạt: DNA cho vòng 1 | 0,80× | 0,70× | 0,60× | 0,55× | 0,50× | 0,45× |
| Tỷ lệ hạt: DNA cho lần thứ 2 Tròn | 0,20× | 0,20× | 0,20× | 0,15× | 0,15× | 0,15× |
Lưu ý: "×" trong bảng biểu thị thể tích của mẫu DNA. Ví dụ, nếu chiều dài chèn của thư viện là 250 bp và thể tích mẫu DNA là 100 μL, thể tích hạt từ được sử dụng trong vòng phân loại đầu tiên là 0,80×100 μL=80 μL; thể tích hạt từ được sử dụng trong vòng phân loại thứ hai là 0,20×100 μL=20 μL.
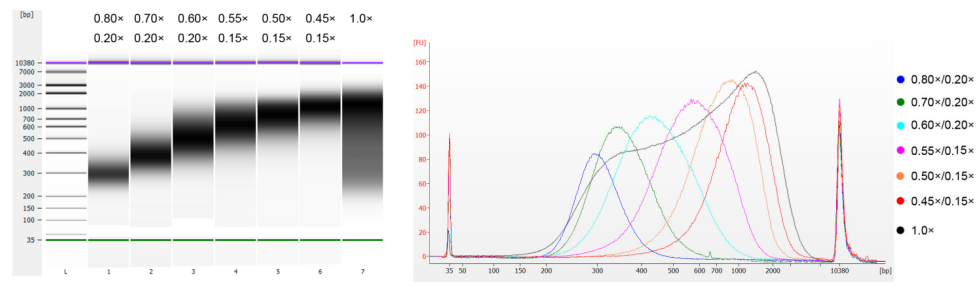
Hình 2. Điện di DNA chip độ nhạy cao Agilent 2100
Ghi chú:
1. Vì sự an toàn và sức khỏe của bạn, vui lòng mặc áo khoác phòng thí nghiệm và găng tay dùng một lần khi thực hiện thí nghiệm.

Trích từ "Tích hợp trình tự cụ thể của họ 1 casposase từ Candidatus Nitrosopumilus koreensis AR1. Nghiên cứu về axit nucleic 2021;49(17):9938-9952. doi:10.1093/nar/gkab725"

Trích từ "Nhiễm trùng gần đây do Wolbachia làm thay đổi cộng đồng vi khuẩn trong quần thể Laodelphax striatellus hoang dã. Hệ vi sinh vật. 2020;8(1):104. Xuất bản ngày 2 tháng 7 năm 2020. doi:10.1186/s40168-020-00878-x"
[1] Wang X, Yuan Q, Zhang W, et al. Tích hợp trình tự cụ thể bởi họ 1 casposase từ Candidatus Nitrosopumilus koreensis AR1. Nghiên cứu về axit nucleic 2021;49(17):9938-9952. doi:10.1093/nar/gkab725(IF:16.971)
[2] Duan XZ, Sun JT, Wang LT, et al. Nhiễm trùng gần đây do Wolbachia làm thay đổi cộng đồng vi khuẩn trong quần thể Laodelphax striatellus hoang dã. Microbiome. 2020;8(1):104. Xuất bản ngày 2 tháng 7 năm 2020. doi:10.1186/s40168-020-00878-x(IF:11.607)
[3] Song B, Almatrafi E, Sang F, et al.Quản lý trầm tích được xử lý bằng Fenton với than sinh học và phân chuồng cừu: Tác động đến đặc điểm tiến hóa của cộng đồng vi khuẩn. J Environ Manage. 2022;316:115218. doi:10.1016/j.jenvman.2022.115218(IF:6.789)
[4] Huang C, Mei Q, Lou L, et al. Viêm loét đại tràng đáp ứng với cấy ghép vi khuẩn đường ruột thông qua điều chỉnh vi khuẩn đường ruột và cân bằng tế bào Th17/Treg. Tế bào. 2022;11(11):1851. Xuất bản ngày 5 tháng 6 năm 2022. doi:10.3390/cells11111851(IF:6.600)
[5] Ghosh S, Yang X, Wang L, Zhang C, Zhao L. Cho ăn prebiotic pha hoạt động làm thay đổi hệ vi khuẩn đường ruột, gây ra sự giảm độc lập với trọng lượng của tình trạng gan nhiễm mỡ và cholesterol huyết thanh ở chuột được cho ăn chế độ nhiều chất béo. Comput Struct Biotechnol J. 2020;19:448-458. Xuất bản ngày 24 tháng 12 năm 2020. doi:10.1016/j.csbj.2020.12.011(IF:6.018)
[6] Gao X, Yu B, Yu J, et al. Hồ sơ phát triển tiêu hóa carbohydrate trong chế độ ăn ở lợn con. Front Microbiol. 2022;13:896660. Xuất bản ngày 29 tháng 4 năm 2022. doi:10.3389/fmicb.2022.896660(IF:5.640)
[7] Li P, Zhang Y, Yan F, Zhou X. Đặc điểm của một loại vi khuẩn thực khuẩn, vB_Kox_ZX8, được phân lập từ Klebsiella oxytoca lâm sàng và tác dụng điều trị của nó đối với vi khuẩn huyết ở chuột. Front Microbiol. 2021;12:763136. Xuất bản ngày 3 tháng 12 năm 2021. doi:10.3389/fmicb.2021.763136(IF:5.640)
[8] Lin Z, Luo P, Huang D, Wu Y, Li F, Liu H. Chiến lược dựa trên đa ô-míc để phân tích độc tính của acrylamide trong mô hình Saccharomyces cerevisiae. Chem Biol Interact. 2021;349:109682. doi:10.1016/j.cbi.2021.109682(IF:5.194)
[9] Sun X, Lv W, Wang Y, và những người khác. Gen Mrgprb2 đóng vai trò trong các phản ứng phản vệ do tiêm Houttuynia cordata gây ra. J Ethnopharmacol. 2022;289:115053. doi:10.1016/j.jep.2022.115053(IF:4.360)
[10] Ma H, Lai B, Zan C, Di X, Zhu X, Wang K. GLO1 góp phần vào khả năng kháng thuốc của Escherichia coli thông qua việc kích thích loại PER của β-Lactamase phổ mở rộng. Nhiễm trùng kháng thuốc. 2022;15:1573-1586. Xuất bản ngày 5 tháng 4 năm 2022. doi:10.2147/IDR.S358578(IF:4.003)
[11] Zhong Y, Zhao W, Tang Z, et al. Phân tích phiên mã so sánh các giai đoạn phát triển khác nhau của buồng trứng ở tôm càng đỏ đầm lầy Procambarus clarkii. BMC Genomics. 2021;22(1):199. Xuất bản ngày 21 tháng 3 năm 2021. doi:10.1186/s12864-021-07537-x(IF:3.969)
[12] Lian C, Yang H, Lan J, et al. Phân tích so sánh bộ gen lục lạp cho thấy mối quan hệ phát sinh loài và biến thể nội loài trong cây thuốc Isodon rubescens. PLoS One. 2022;17(4):e0266546. Xuất bản ngày 6 tháng 4 năm 2022. doi:10.1371/journal.pone.0266546(IF:3.240)
[13] Diao G, Huang J, Zheng X, và cộng sự. Prostaglandin E2 đóng vai trò kép trong việc điều chỉnh sự di chuyển của các tế bào dendritic. Int J Mol Med. 2021;47(1):207-218. doi:10.3892/ijmm.2020.4801(IF:3.098)
[14] Bing XL, Zhao DS, Peng CW, Huang HJ, Hong XY. Điểm tương đồng và biến thể không gian của cộng đồng vi khuẩn và nấm trong quần thể rầy nâu hại lúa (Hemiptera: Delphacidae). Insect Sci. 2020;27(5):947-963. doi:10.1111/1744-7917.12782(IF:2.791)
[15] Li X, Zhou S, Zhang J, Zhou Z, Xiong Q. Những thay đổi theo hướng của cộng đồng vi khuẩn đường ruột ở ấu trùng ruồi lính đen (Hermetia illucens). Động vật (Basel). 2021;11(12):3475. Xuất bản ngày 6 tháng 12 năm 2021. doi:10.3390/ani11123475(IF:2.752)
[16] Yang J, Peng Y, Kong W. Xác định và giải trình tự bộ gen hoàn chỉnh của virus bí ẩn dâu tằm 1. Arch Virol. 2022;167(2):687-690. doi:10.1007/s00705-021-05350-1(IF:2.574)
[17] Chang Y, Xia X, Sui L, et al. Sự xâm chiếm nội sinh của nấm gây bệnh cho côn trùng làm tăng khả năng kháng bệnh của cây bằng cách thay đổi cộng đồng vi khuẩn nội sinh. J Basic Microbiol. 2021;61(12):1098-1112. doi:10.1002/jobm.202100494(IF:2.281)
[18] Ding CY, Ma YM, Li B, et al. Xác định và phân tích chức năng của các gen biểu hiện khác biệt ở Myzus persicae (Hemiptera: Aphididae) để đáp ứng với Trans-anethole. J Insect Sci. 2022;22(1):3. doi:10.1093/jisesa/ieab094(IF:1.857)
Thanh toán & Bảo mật
Thông tin thanh toán của bạn được xử lý an toàn. Chúng tôi không lưu trữ chi tiết thẻ tín dụng cũng như không có quyền truy cập vào thông tin thẻ tín dụng của bạn.
Cuộc điều tra
Bạn cũng có thể thích
Câu hỏi thường gặp
Sản phẩm chỉ dành cho mục đích nghiên cứu và không dùng để điều trị hoặc chẩn đoán ở người hoặc động vật. Sản phẩm và nội dung được bảo vệ bởi các bằng sáng chế, nhãn hiệu và bản quyền thuộc sở hữu của
Một số ứng dụng có thể yêu cầu thêm quyền sở hữu trí tuệ của bên thứ ba.