

Sự miêu tả
Bộ phát hiện Mycoplasma GMyc-PCR chủ yếu sử dụng phương pháp PCR để phát hiện nhiễm trùng Mycoplasma của nhiều vật liệu sinh học khác nhau (như nuôi cấy tế bào, dịch tiết động vật thí nghiệm, huyết thanh động vật, v.v.). Nó kết hợp một số ưu điểm: nhạy cảm, đặc hiệu, nhanh chóng và có thể phát hiện trực tiếp bằng dịch nuôi cấy tế bào. Sản phẩm này phát hiện mycoplasma trong các vật liệu sinh học như tế bào nuôi cấy bằng phương pháp PCR. Các đoạn mồi được sử dụng được thiết kế theo vùng bảo tồn của trình tự rRNA 16S-23S của mycoplasma và chỉ khuếch đại cụ thể DNA mycoplasma, với độ nhạy và độ đặc hiệu phát hiện cao. Phân tích khuếch đại PCR và điện di chỉ mất vài giờ, thao tác thuận tiện và đơn giản.
Nuôi cấy tế bào là một thí nghiệm phổ biến trong nghiên cứu khoa học sự sống. Không giống như các phương pháp thí nghiệm thường dùng khác, nuôi cấy tế bào là một quá trình liên tục động và các tế bào thường phản ứng với các lỗi thao tác hoặc chất gây ô nhiễm thường biểu hiện trạng thái tế bào hoặc hình dạng môi trường bất thường. Nếu bị nhiễm mycoplasma, hình thái tế bào không có thay đổi rõ ràng và dễ bị bỏ qua. Thường không phát hiện ra cho đến khi ô nhiễm trở nên rất nghiêm trọng. Có thể có hàng trăm mycoplasma trên màng tế bào bị ô nhiễm, những mycoplasma này cạnh tranh chất dinh dưỡng và giải phóng các chất chuyển hóa độc hại, ảnh hưởng nghiêm trọng đến kết quả thí nghiệm.
Các nghiên cứu đã chỉ ra rằng có ít nhất 20 loại mycoplasma có thể gây nhiễm tế bào, trong đó phổ biến nhất là: Mycoplasma đường uống (M. orale), Mycoplasma arginine (M. arginini), Mycoplasma hyorhinis (M. hyorhinis), Mycoplasma fermentum (M. lên men), Mycoplasma hominis (M. hominis), Mycoplasma salivarius (M. salivarium), Mycoplasma phổi (M. pulmonis) và Mycoplasma lê (M. pirum). Tỷ lệ nhiễm mycoplasma của tế bào nuôi cấy dao động từ 4% đến 92%. Các nguồn nhiễm bao gồm môi trường làm việc, bản thân người vận hành (một số mycoplasma là hệ vi khuẩn bình thường của cơ thể người), môi trường nuôi cấy, huyết thanh, nhiễm chéo tế bào, thiết bị thí nghiệm và nhiễm bẩn đã sử dụng của mô hoặc cơ quan ban đầu mà từ đó tế bào được chuẩn bị.
Xác định nguyên nhân cơ bản của các vấn đề trong quá trình nuôi cấy tế bào là một nhiệm vụ khó khăn và tốn thời gian, trong đó bất kỳ thay đổi đột ngột nào cũng cần được nghi ngờ và cần có các biện pháp kiểm tra tốt và kiểm tra thường xuyên để phát hiện nhiễm mycoplasma. Có nhiều phương pháp để phát hiện mycoplasma, chẳng hạn như nuôi cấy trực tiếp, nhuộm huỳnh quang DNA, ELISA và phương pháp PCR.
Tính năng
- Các đoạn mồi được sử dụng được thiết kế theo vùng bảo tồn của trình tự rRNA 16S-23S của Mycoplasma
- Chỉ có DNA mycoplasma được khuếch đại đặc hiệu
- Độ nhạy và độ đặc hiệu tuyệt vời
- Có thể phát hiện hơn 20 loài mycoplasma
Ứng dụng
- Phát hiện Mycoplasma
Thành phần
| Số thành phần | Tên | 40601ES10 (10 xét nghiệm) | 40601ES20 (20 xét nghiệm) |
| 40601-A | Hỗn hợp PCR GMyc-1st | 250 µL | 2×250 µL |
| 40601-B | Hỗn hợp PCR GMyc-2nd | 250 µL | 2×250 µL |
| 40601-C | Mẫu kiểm soát tích cực N | 20 µL | 20 µL |
[Lưu ý] 1. Khi không sử dụng trong thời gian dài, có thể bảo quản đông lạnh ở nhiệt độ -85~-65℃.
2. Phản ứng PCR cực kỳ nhạy.Để tránh kết quả dương tính giả, một mẫu chứng dương tính sẽ được thêm vào cuối khi thêm mẫu.
Kho
Sản phẩm này có thể được bảo quản ở nhiệt độ -25~-15℃ trong 18 tháng. Nếu không sử dụng trong thời gian dài, vui lòng để xa ánh sáng.
Các con số
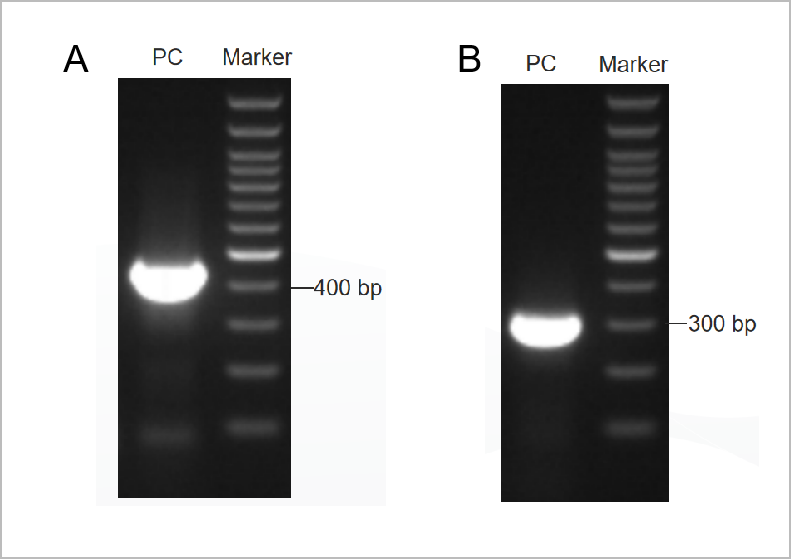
Hình 1. Kết quả điện di của bộ xét nghiệm PCR Mycoplama.
Điện di vòng đầu tiên được thể hiện ở Hình 1A và điện di vòng thứ hai được thể hiện ở Hình 1B. (M: dấu hiệu 1kb, 1-2: đối chứng dương, 3: đối chứng âm)
[1] Rao XS, Cong XX, Gao XK, et al. Sự phosphoryl hóa trung gian bởi AMPK làm tăng sự tự ức chế của TBC1D17 để thúc đẩy sự hấp thụ glucose phụ thuộc Rab5. Cell Death Differ. 2021;28(12):3214-3234. doi:10.1038/s41418-021-00809-9(IF:15.828)
[2] Guo F, Li L, Li J, et al. Trình tự đa ô-míc tế bào đơn của phôi chuột giai đoạn đầu và tế bào gốc phôi. Cell Res. 2017;27(8):967-988. doi:10.1038/cr.2017.82(IF:15.606)
[3] Hao Y, He B, Wu L, et al. Sự chuyển vị hạt nhân của p85β thúc đẩy quá trình hình thành khối u của ung thư đột biến miền xoắn ốc PIK3CA. Nat Commun. 2022;13(1):1974. Xuất bản ngày 13 tháng 4 năm 2022. doi:10.1038/s41467-022-29585-x(IF:14.919)
[4] Shu X, Liu M, Lu Z, et al. Bản đồ toàn bộ hệ gen cho thấy deoxyuridine được làm giàu trong DNA trung tâm của con người. Nat Chem Biol. 2018;14(7):680-687. doi:10.1038/s41589-018-0065-9(IF:13.843)
[5] Li X, Xiong X, Wang K, et al. Bản đồ toàn bộ transcriptome cho thấy methylome N(1)-methyladenosine có thể đảo ngược và động. Nat Chem Biol. 2016;12(5):311-316. doi:10.1038/nchembio.2040(IF:12.709)
[6] Sun L, Yang X, Huang X, et al. 2-Hydroxylation of Fatty Acids Represses Rummigenesis and Metastasis of Rummistance through the YAP Transcriptional Axis. Cancer Res. 2021;81(2):289-302. doi:10.1158/0008-5472.CAN-20-1517(IF:12.701)
[7] Sun Z, Zhang Z, Wang QQ, Liu JL. Sự kết hợp bất hoạt của CTPS1 và ATR gây tử vong tổng hợp cho các tế bào ung thư biểu hiện quá mức MYC. Cancer Res. 2022;82(6):1013-1024. doi:10.1158/0008-5472.CAN-21-1707(IF:12.701)
[8] Song J, Zhuang Y, Zhu C, et al. Các vai trò khác nhau của PUS10 ở người trong quá trình xử lý miRNA và pseudouridylation tRNA. Nat Chem Biol. 2020;16(2):160-169. doi:10.1038/s41589-019-0420-5(IF:12.154)
[9] He B, Pan H, Zheng F, et al. RNA dài không mã hóa LINC00930 thúc đẩy quá trình phân giải tế bào khối u và tăng sinh tế bào do PFKFB3 trung gian trong ung thư vòm họng. J Exp Clin Cancer Res. 2022;41(1):77. Xuất bản ngày 24 tháng 2 năm 2022. doi:10.1186/s13046-022-02282-9(IF:11.161)
[10] Tang B, Liu BH, Liu ZY, Luo MY, Shi XH, Pang DW. Các chấm lượng tử với lớp phủ lưỡng tính lưỡng tính nhỏ gọn. Giao diện vật liệu ứng dụng ACS. 2022;14(24):28097-28104. doi:10.1021/acsami.2c04438(IF:9.229)
[11] Huang C, Zhang Z, Chen L, và những người khác.Acetyl hóa trong miền đầu cuối N và C của Src điều chỉnh các vai trò riêng biệt của quá trình sinh khối u trung gian bởi STAT3. Cancer Res. 2018;78(11):2825-2838. doi:10.1158/0008-5472.CAN-17-2314(IF:9.130)
[12] Wu X, Yu M, Zhang Z, et al. DDB2 điều chỉnh quá trình sao chép DNA thông qua quá trình phân hủy CDT2 độc lập với PCNA. Cell Biosci. 2021;11(1):34. Xuất bản ngày 8 tháng 2 năm 2021. doi:10.1186/s13578-021-00540-5(IF:7.133)
[13] Wang J, Zhang Y, Liu X, Liu H. Tối ưu hóa liệu pháp thích ứng dựa trên khả năng tiếp cận với nhóm dân số kháng khối u. Ung thư (Basel). 2021;13(21):5262. Xuất bản ngày 20 tháng 10 năm 2021. doi:10.3390/cancers13215262(IF:6.639)
[14] Feng W, Liu R, Xie X, et al. SUMOylation của α-tubulin là một sửa đổi mới điều chỉnh động lực học của vi ống. J Mol Cell Biol. 2021;13(2):91-103. doi:10.1093/jmcb/mjaa076(IF:6.216)
[15] Yu M, Hu X, Yan J, Wang Y, Lu F, Chang J. Chất ức chế RIOK2 NSC139021 phát huy tác dụng chống khối u trên u nguyên bào thần kinh đệm thông qua việc gây ra sự bắt giữ chu kỳ tế bào và apoptosis do Skp2 làm trung gian. Y sinh học. 2021;9(9):1244. Xuất bản ngày 17 tháng 9 năm 2021. doi:10.3390/biomedicines9091244(IF:6.081)
[16] Ren S, Cai Y, Hu S, et al. Berberine phát huy hoạt động chống khối u trong u lympho tế bào B lớn lan tỏa bằng cách điều chỉnh trục c-myc/CD47. Biochem Pharmacol. 2021;188:114576. doi:10.1016/j.bcp.2021.114576(IF:5.858)
[17] Wen F, Sun X, Sun C, et al. TAGLN bị điều hòa giảm bởi sự thoái hóa proteasomal do TRAF6 làm trung gian trong tế bào ung thư tuyến tiền liệt. Mol Cancer Res. 2021;19(7):1113-1122. doi:10.1158/1541-7786.MCR-20-0513(IF:5.852)
[18] Tang B, Sun EZ, Zhang ZL, et al. Miền cholesterol được cô lập bằng sphingomyelin tuyển dụng protein liên kết với formin 17 để thắt chặt các hố phủ clathrin trong quá trình xâm nhập của vi-rút cúm. J Virol. 2022;96(5):e0181321. doi:10.1128/JVI.01813-21(IF:5.103)
[19] Hu J, Ren W, Qiu W, et al. Tạo ra dòng tế bào gốc đa năng cảm ứng (XDCMHi001-A) từ bệnh nhân viêm cột sống dính khớp có đột biến JAK2. Stem Cell Res. 2020;45:101788. doi:10.1016/j.scr.2020.101788(IF:4.495)
[20] Xiao S, Yao X, Ye J, Tian X, Yin Z, Zhou L. Biến đổi biểu sinh tạo điều kiện cho biểu hiện bất thường của proline synthase PYCR1 trong ung thư dạ dày [được xuất bản trực tuyến trước khi in, ngày 30 tháng 5 năm 2022]. Biochim Biophys Acta Gene Regul Mech. 2022;1865(6):194829. doi:10.1016/j.bbagrm.2022.194829(IF:4.490)
[21] Wang J, Zhang Y, Liu X, Liu H. Liệu phương pháp điều trị định kỳ cố định có hiệu quả đối với hệ thống khối u mà không có thông tin đầy đủ không?. Cancer Manag Res. 2021;13:8915-8928. Xuất bản ngày 30 tháng 11 năm 2021. doi:10.2147/CMAR.S339787(IF:3.989)
[22] Yang X, Ren S, Rehman ZU, et al. Đặc điểm phân tử, biểu hiện và nhận dạng chức năng của TANK-binding kinase 1 (TBK1) ở bò (Bos taurus) và dê (Capra hircus). Dev Comp Immunol. 2022;133:104444. doi:10.1016/j.dci.2022.104444(IF:3.636)
[23] Zheng D, Chang X, Liu Y, et al. 2-Methoxy-5((3,4,5-trimethosyphenyl)seleninyl) phenol đảo ngược quá trình di cư và xâm lấn tế bào do EGF gây ra thông qua việc điều hòa giảm MDM2 trong các dòng tế bào ung thư vú. Cancer Biol Ther. 2019;20(4):513-523. doi:10.1080/15384047.2018.1537578(IF:3.373)
[24] Xu F, Zhang S, Liu Z, et al. TEX9 và eIF3b có chức năng hiệp đồng thúc đẩy sự tiến triển của ung thư biểu mô tế bào vảy thực quản. BMC Cancer. 2019;19(1):875. Xuất bản ngày 3 tháng 9 năm 2019. doi:10.1186/s12885-019-6071-9(IF:2.933)
[25] Pan H, Sun L, Wang W, et al. RNA không mã hóa dài trong huyết thanh LOC553103 là dấu ấn sinh học chẩn đoán và tiên lượng không đặc hiệu cho các loại ung thư phổ biến ở người. Clin Chim Acta. 2020;508:69-76. doi:10.1016/j.cca.2020.05.017(IF:2.615)
[26] Li N, Lin SM, Li Y, Sun J, Zhang L, Chen M.Dòng tế bào gốc đa năng cảm ứng (GZHMCi004-A) có nguồn gốc từ thai nhi có đột biến G380R dị hợp tử ở gen FGFR3 gây ra chứng loạn sản sụn. Stem Cell Res. 2021;53:102322. doi:10.1016/j.scr.2021.102322(IF:2.020)
[27] Luo Q, Wei C, Long Y, et al. Tạo ra dòng tế bào gốc phôi người loại bỏ gen ELTD1 bằng hệ thống iCRISPR/Cas9. Stem Cell Res. 2021;53:102350. doi:10.1016/j.scr.2021.102350(IF:2.020)
[28] Liu YQ, Ling TW, Wang HY, Yang YH, Song WJ, Wang TC. Tạo ra dòng tế bào gốc đa năng cảm ứng không tích hợp (LZUSHI001-A) từ bệnh nhân động kinh có đột biến DGKG. Stem Cell Res. 2022;61:102768. doi:10.1016/j.scr.2022.102768(IF:2.020)
[29] Chen M, Lin SM, Li N, Li Y, Li Y, Zhang L. Dòng tế bào gốc đa năng cảm ứng (GZHMCi003-A) có nguồn gốc từ thai nhi bị mất đoạn exon 3 dị hợp tử trong gen RUNX2 gây ra chứng loạn sản xương cleidoccraial. Stem Cell Res. 2021;51:102166. doi:10.1016/j.scr.2021.102166(IF:2.020)
[30] Xu Y, Wang X, Qiu T, et al. Tạo ra dòng tế bào gốc đa năng cảm ứng (FDCHI007-A) có nguồn gốc từ bệnh nhân mắc bệnh não phát triển và động kinh loại 31 mang gen c.545C dị hợp tử > Đột biến trong gen DNM1. Stem Cell Res. 2022;60:102709. doi:10.1016/j.scr.2022.102709(IF:2.020)
[31] Fan T, He J, Wang Y, Yu J, Sun W. Tạo ra dòng tế bào gốc đa năng cảm ứng (FDCHi006-A) từ một bé gái 7 tuổi bị dậy thì sớm trung ương. Stem Cell Res. 2021;56:102542. doi:10.1016/j.scr.2021.102542(IF:2.020)
[32] Gong X, Zheng Z, Yang T, Zheng H, Xiao X, Jia N. Tạo ra dòng iPSC được hiệu chỉnh gen đồng hợp tử (OGHFUi001-A-1) từ bệnh nhân mắc bệnh não động kinh ở trẻ sơ sinh loại 1 (EIEE1) có đột biến R330L đồng hợp tử ở gen ARX. Stem Cell Res. 2022;60:102693. doi:10.1016/j.scr.2022.102693(IF:2.020)
[33] Jia N, Gong X, Chen J, et al. Tạo ra dòng tế bào gốc đa năng cảm ứng (OGHFUi001-A) từ bệnh não động kinh trẻ sơ sinh loại 1 có đột biến ARX. Stem Cell Res. 2021;53:102367. doi:10.1016/j.scr.2021.102367(IF:2.020)
[34] Zhu W, Zhou Y, Wang Q, et al. Tạo ra dòng tế bào gốc đa năng cảm ứng của con người (iPSC) từ nguyên bào sợi da của bệnh nhân mang đột biến E363Q trong gen PSEN1. Stem Cell Res. 2022;61:102769. doi:10.1016/j.scr.2022.102769(IF:2.020)
[35] Luo F, Long K, Li X, et al. Thiếu hụt LRRC8A làm giảm hoại tử do thiếu oxy ở tế bào 3T3-L1. Biosci Biotechnol Biochem. 2020;84(6):1139-1145. doi:10.1080/09168451.2020.1730689(IF:1.516)
Thanh toán & Bảo mật
Thông tin thanh toán của bạn được xử lý an toàn. Chúng tôi không lưu trữ chi tiết thẻ tín dụng cũng như không có quyền truy cập vào thông tin thẻ tín dụng của bạn.
Cuộc điều tra
Bạn cũng có thể thích
Câu hỏi thường gặp
Sản phẩm chỉ dành cho mục đích nghiên cứu và không dùng để điều trị hoặc chẩn đoán ở người hoặc động vật. Sản phẩm và nội dung được bảo vệ bởi các bằng sáng chế, nhãn hiệu và bản quyền thuộc sở hữu của
Một số ứng dụng có thể yêu cầu thêm quyền sở hữu trí tuệ của bên thứ ba.