


Beschreibung
Hieff Trans™ Das liposomale Transfektionsreagenz ist ein vielseitiges Liposomen-Transfektionsreagenz, geeignet für die DNA-, RNA- und Oligonukleotid-Transfektion und bietet eine hohe Transfektionseffizienz für die meisten eukaryotischen Zellen. Seine einzigartige Formel ermöglicht die direkte Zugabe zum Medium. Serum beeinträchtigt die Transfektionseffizienz nicht, wodurch Zellschäden durch Serumentfernung reduziert werden. Der Nukleinsäure-Hieff Trans™-Komplex muss nach der Transfektion weder entfernt noch durch frisches Medium ersetzt werden; er kann auch nach 4–6 Stunden entfernt werden.
Hieff Trans™ wird in steriler flüssiger Form geliefert. Normalerweise werden für die Transfektion in 24-Well-Platten jeweils etwa 1,5 μL, 1 ml Hieff Trans™ kann etwa 660 Transfektionen durchführen; bei einer 6-Well-Platte etwa 6 μl jedes Mal, 1 ml Hieff Trans™ kann etwa 660 Transfektionen durchführen. 160 Transfektionen.
Besonderheit
- Außergewöhnliche Effizienz: Überlegene Transfektionsleistung in einem breiten Spektrum von Zelllinien, einschließlich transienter und stabiler Transfektion
- Breite Anpassungsfähigkeit: Hervorragende Transfektionseffizienz in einer Vielzahl von Zelllinien und hohe Expression rekombinanter Proteine
- Geringe Toxizität: Die Aktivitäten der verschiedenen Zellen werden durch Transfektionsreagenzien nahezu nicht beeinflusst.
YEASEN - Einfache Bedienung: Nachgewiesene Wirksamkeit in Gegenwart von Serum – macht einen Medienwechsel nach der Transfektion überflüssig
- Kostengünstig: Wettbewerbsfähiger Transfektionseffekt zu günstigeren Preisen
Anwendung
- Zelltransfektion
Spezifikation
| Bilden | Flüssig |
| Serumkompatibel | Ja |
| Zelltyp | Etablierte Zelllinien |
| Probentyp | Plasmid-DNA, synthetische siRNA |
| Transfektionstechnik | Lipidbasierte Transfektion |
Komponenten
| Komponenten Nr. | Name | 40802ES02 | 40802ES03 | 40802ES08 |
| 40802 | Hieff Trans™ Liposomales Transfektionsreagenz | 0,5 ml | 1 ml | 5×1 ml |
Versand und Lagerung
Das Produkt wird mit Kühlakkus geliefert und ist bei 2-8ºC ein Jahr lagerfähig. Nicht einfrieren!
Zahlen

Zitiert nach: Communications Biology Band 5, Artikelnummer: 1296 (2022)
- Demonstration des Transfektionseffekts
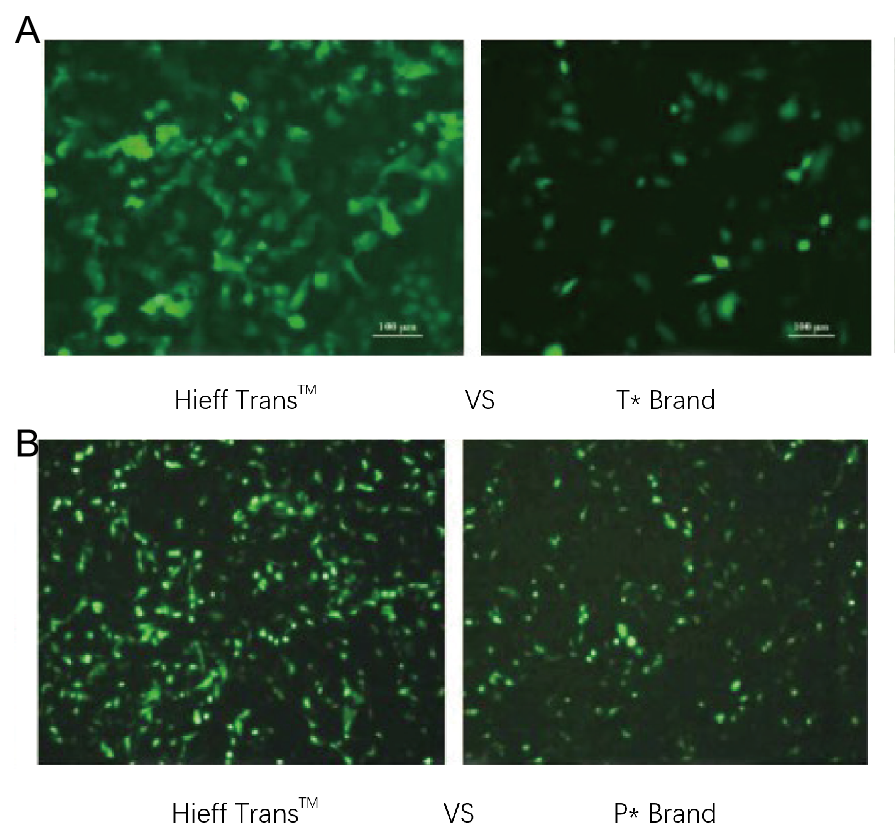
Abbildung 1. Das Hieff Trans™ Liposome Transfection Reagent übertrifft das Transfektionsreagenz der Konkurrenzmarke (Marke T* in Abbildung 1a, Marke P* in Abbildung 1b).
Jedes Reagenz wurde zur Transfektion der Zielzelllinie (Hela-Zelllinie in Abbildung 1a, DF-1-Zelllinie in Abbildung 1b) in einem 96-Well-Format verwendet. Die GFP-Expression wurde 48 Stunden nach der Transfektion analysiert. Das Hieff Trans™ Liposomen-Transfektionsreagenz zeigte eine höhere GFP-Transfektionseffizienz als Konkurrenzprodukte.
- Validierte Zelllinien
| Produkt Name | Hieff TransTMLiposomal Transfektion Reagens | |||||
| Validiert Zelle Linien | 293T | Calu 1 | HEK293 | HO1980 | N2A | TS |
| 293F | CHO-K1 | HEK293T | HUVEC | NCI-H1975 | U-87 | |
| 293 Fuß | COS-7 | Hela | MCF10A | NIH-3T3 | Vero | |
| 3T3 | DF-1 | Hep2C | MCF-7 | Neuro-2a | WEHI | |
| A549 | H1299 | Hep3B | MDA-MB-231 | PC-12 | WRL-68 | |
| BV-2 | H520 | Hepa1-6 | MDA-MB-231-LM2-4175 | Raw264.7 | ||
| B50 | HaCaT | HepG2 | MDCK | SGC-7901 | ||
| C2C12 | HCT116 | HK2 | MEF | T47D | … | |
Abbildung 2. Liste der erfolgreich mit Hieff Trans™ transfizierten Zelllinien Liposomales Transfektionsreagenz (wird ständig aktualisiert).

Zitiert aus „Optogenetische Kontrolle der RNA-Funktion und des RNA-Stoffwechsels mithilfe konstruierter lichtschaltbarer RNA-bindender Proteine. Nat Biotechnol. 3. Januar 2022. doi: 10.1038/s41587-021-01112-1"

Zitiert aus „Strukturelle Einblicke in die Rekrutierung des BRCA1-BARD1-Komplexes zu beschädigtem Chromatin. Mol Cell. 2021 Jul 1;81(13):2765-2777.e6. doi: 10.1016/j.molcel.2021.05.010.“
Zitiert aus „Die UBQLN2-HSP70-Achse reduziert Poly-Gly-Ala-Aggregate und lindert Verhaltensdefekte im C9ORF72-Tiermodell. Neuron. 16. Juni 2021;109(12):1949-1962.e6. doi: 10.1016/j.neuron.2021.04.023.“
Hieff Trans™ Liposomales Transfektionsreagenz Häufig gestellte Fragen
(1) F: Kann bei der Herstellung des Nukleinsäure-Transfektionsreagenzkomplexes Serum vorhanden sein?
A: Das Vorhandensein von Serum beeinflusst die Bildung von Liposomen. Es wird empfohlen, bei der Herstellung von Nukleinsäure-Transfektionsreagenzkomplexen serumfreies Medium (normalerweise MEM-Medium) zu verwenden.
(2) F: Kann das Transfektionsreagenz eingefroren werden?
A: Nein. Dieses Reagenz muss bei 2–8 °C gelagert werden. Achten Sie darauf, den Deckel nicht über einen längeren Zeitraum wiederholt zu öffnen, da ein langfristiges Öffnen des Deckels zu einer Oxidation der Liposomen führt und die Transfektionseffizienz beeinträchtigt.
(3) F: Worauf muss ich bei der Verwendung des Hieff Trans™ Liposome Nucleic Acid Transfection Reagent achten?
A: 1) Während des Transfektionsvorgangs ist es besser, wenn die Zellkonfluenz 80 % – 95 % erreicht, und die spezifische Beschichtungsdichte wird entsprechend der Situation der Zellen bestimmt;
2) Die Verwendung hochreiner DNA trägt dazu bei, eine höhere Transfektionseffizienz zu erzielen.
3) DNA und Transfektionsreagenzien müssen bei der Herstellung von Transfektionskomplexen mit serumfreiem Medium verdünnt werden;
4) Während der Transfektion dürfen dem Medium keine Antibiotika zugesetzt werden.
5) Die DNA-Konzentration und die Menge des kationischen Liposomenreagenzes sollten für den ersten Einsatz optimiert werden, um eine maximale Transfektionseffizienz zu erzielen. Das Verhältnis von DNA zu Transfektionsreagenz beträgt Im Allgemeinen wird ein Verhältnis von 1:2–1:3 empfohlen.
(4) F: Muss die Transfektion nach der Transfektion beendet werden?
A: Nicht erforderlich. Liposomenkomplexe sind 6 Stunden stabil. Wird das Zellmedium vor der Transfektion nicht gewechselt, ist nach 4 bis 6 Stunden ein Wechsel auf ein neues Medium erforderlich, um die für ein normales Zellwachstum notwendigen Nährstoffe sicherzustellen. Wurde das Medium jedoch vor der Transfektion gewechselt, ist ein Wechsel nach der Liposomentransfektion nicht erforderlich.
(5) F: Worauf muss ich achten, wenn ich die Transfektionseffizienz verbessern möchte?
A: a: Die Zelldichte zum Zeitpunkt der Transfektion beträgt 90 %–95 %.
b: Verwenden Sie während der Transfektion serumfreies MEM-Medium für Nukleinsäure- und Liposomenverdünnungen.
c: Das Medium kann 4-6 Stunden nach der Transfektion gewechselt werden.
(6) F: Ist eine Kotransfektion von DNA und siRNA möglich? Wie ist die Wirkung?
A: Eine Kotransfektion ist möglich, es wird jedoch empfohlen, die Transfektion separat durchzuführen. Die DNA-Transfektion sollte 6 Stunden nach der siRNA-Transfektion erfolgen. Bei gleichzeitiger Durchführung verringert sich die Effizienz der siRNA-Transfektion.
(7) F: Kann das Transfektionsreagenz für die lentivirale Verpackungstransfektion verwendet werden?
A: Eine lentivirale Verpackung ist möglich, aber die Effizienz der lentiviralen Verpackung hängt nicht unbedingt mit der Effizienz der Transfektion zusammen, sondern auch mit der Auswahl der Verpackungsplasmide und dem Verhältnis zwischen den Plasmiden.
(8) F: Kann das Hieff Trans™ Liposome Nucleic Acid Transfection Reagent zur Transfektion von Suspensionszellen verwendet werden?
A: Das Hieff Trans™ Liposomen-Nukleinsäure-Transfektionsreagenz kann für die Transfektion von Suspensionszellen verwendet werden. Einzelheiten finden Sie im Protokoll. Darüber hinaus haben wir ein Transfektionsreagenz speziell für Suspensionszellen auf den Markt gebracht (Kat.-Nr. 40805, Liposomen-Nukleinsäure-Transfektionsreagenz für Suspensionszellen).
[1] Liu R, Yang J, Yao J, Zhao Z, He W, Su N, Zhang Z, Zhang C, Zhang Z, Cai H, Zhu L, Zhao Y, Quan S, Chen X, Yang Y. Opto-genetische Kontrolle der RNA-Funktion und des RNA-Stoffwechsels mithilfe künstlich hergestellter lichtschaltbarer RNA-bindender Proteine. Nat Biotechnol. 3. Januar 2022. doi: 10.1038/s41587-021-01112-1. Epub ahead of print. PMID: 34980910. (IF:54.908)
[2] Zhou J, Chen P, Wang H, Liu H, Li Y, Zhang Y, Wu Y, Paek C, Sun Z, Lei J, Yin L. Cas12a-Varianten, die durch strenge PAM-Erkennung für einen geringeren genomweiten Off-Target-Effekt entwickelt wurden. Mol Ther. 5. Januar 2022;30(1):244-255. doi: 10.1016/j.ymthe.2021.10.010. Epub 20. Oktober 2021. PMID: 34687846; PMCID: PMC8753454. (IF:11.454)
[3] Chen S, Cao X, Zhang J, Wu W, Zhang B, Zhao F. circVAMP3 treibt die CAPRIN1-Phasentrennung voran und hemmt das hepatozelluläre Karzinom durch Unterdrückung der c-Myc-Translation. Adv Sci (Weinh).2022 März;9(8):e2103817. doi: 10.1002/advs.202103817. Epub 2022 Jan 24. PMID: 35072355; PMCID: PMC8922094. (IF:16.808)
[4] Zhang Y, Yu X, Sun R, Min J, Tang X, Lin Z, Xie S, Li X, Lu S, Tian Z, Gu C, Teng L, Yang Y. Der Spleißfaktor Arginin/Serin-reich 8 fördert die maligne Erkrankung des multiplen Myeloms und Knochenläsionen durch alternatives Spleißen von CACYBP und exosomenbasierte zelluläre Kommunikation. Clin Transl Med. 2022 Feb;12(2):e684. doi: 10.1002/ctm2.684. PMID: 35184390. (IF:11.492)
[5] Tang J Exp Clin Cancer Res. 8. März 2022;41(1):85. doi: 10.1186/s13046-022-02276-7. PMID: 35260179. (IF:11.161)
[6] Hua Z, Wei R, Guo M, Lin Z, Yu X, Li X, Gu C, Yang Y. YTHDF2 fördert die Proliferation multipler Myelomzellen über die STAT5A/MAP2K2/p-ERK-Achse. Oncogene. 2022 März;41(10):1482-1491. doi: 10.1038/s41388-022-02191-3. Epub 2022 Jan 24. PMID: 35075244. (IF:9.867)
[7] Liang Y, Lu Q, Li W, Zhang D, Zhang F, Zou Q, Chen L, Tong Y, Liu M, Wang S, Li W, Ren X, Xu P, Yang Z, Dong S, Zhang B, Huang Y, Li D, Wang H, Yu W. Reaktivierung von Tumorsuppressoren bei Brustkrebs durch Enhancer-Switching über das NamiRNA-Netzwerk. Nucleic Acids Res. 2021 Sep 7;49(15):8556-8572. doi: 10.1093/nar/gkab626. PMID: 34329471; PMCID: PMC8421228. (WENN:16.9)
[8] Dai L, Dai Y, Han J, Huang Y, Wang L, Huang J, Zhou Z. Strukturelle Einblicke in die Rekrutierung des BRCA1-BARD1-Komplexes zu geschädigtem Chromatin. Mol Cell. 2021 Jul 1;81(13):2765-2777.e6. doi: 10.1016/j.molcel.2021.05.010. Epub 2021 Jun 7. PMID: 34102105. (IF:17.97)
[9] Zhang K, Wang A, Zhong K, Qi S, Wei C, Shu X, Tu WY, Xu W, Xia C, Xiao Y, Chen A, Bai L, Zhang J, Luo B, Wang W, Shen C. Die UBQLN2-HSP70-Achse reduziert Poly-Gly-Ala-Aggregate und lindert Verhaltensdefekte im C9ORF72-Tiermodell. Neuron. 16. Juni 2021;109(12):1949-1962.e6. doi: 10.1016/j.neuron.2021.04.023. Epub 14. Mai 2021. PMID: 33991504. (IF:17.17)
[10] Liang Y, Lu Q, Li W, Zhang D, Zhang F, Zou Q, Chen L, Tong Y, Liu M, Wang S, Li W, Ren X, Xu P, Yang Z, Dong S, Zhang B, Huang Y, Li D, Wang H, Yu W. Reaktivierung von Tumorsuppressoren bei Brustkrebs durch Enhancer-Switching über das NamiRNA-Netzwerk. Nucleic Acids Res. 2021 Sep 7;49(15):8556-8572. doi: 10.1093/nar/gkab626. PMID: 34329471; PMCID: PMC8421228. (IF:16.9)
[11] Li T, Chen X, Qian Y, Shao J, Li X, Liu S, Zhu L, Zhao Y, Ye H, Yang Y. Ein synthetisches BRET-basiertes optogenetisches Gerät für die pulsierende Transgenexpression, das die Glukosehomöostase bei Mäusen ermöglicht. Nat Commun. 2021 Jan 27;12(1):615. doi: 10.1038/s41467-021-20913-1. PMID: 33504786; PMCID: PMC7840992. (IF:14.92)
[12] Pan Y, He X, Li C, Li Y, Li W, Zhang H, Wang Y, Zhou G, Yang J, Li J, Qu J, Wang H, Gao Z, Shen Y, Li T, Hu H, Ma H. Neuronale Aktivität rekrutiert die CRTC1/CREB-Achse, um die transkriptionsabhängige Autophagie zur Aufrechterhaltung der späten LTD-Phase voranzutreiben. Cell Rep. 20. Juli 2021;36(3):109398. doi: 10.1016/j.celrep.2021.109398. PMID: 34289350. (IF:9.42)
[13] Liu H, Xing R, Ou Z, Zhao J, Hong G, Zhao TJ, Han Y, Chen Y. Der G-Protein-gekoppelte Rezeptor GPR17 hemmt die Gliomentwicklung durch Erhöhung der durch den Polycomb-Repressivkomplex 1 vermittelten ROS-Produktion. Cell Death Dis. 2021 Jun 12;12(6):610. doi: 10.1038/s41419-021-03897-0. PMID: 34120140; PMCID: PMC8197764. (IF:8.463)
[14] Fan Y, Wang J, Jin W, Sun Y, Xu Y, Wang Y, Liang X, Su D. CircNR3C2 fördert die HRD1-vermittelte tumorsuppressive Wirkung durch Aufsaugen von miR-513a-3p bei dreifach negativem Brustkrebs. Mol Cancer. 2021 Feb 2;20(1):25. doi: 10.1186/s12943-021-01321-x.PMID: 33530981; PMCID: PMC7851937. (IF:27.403)
[15] Gu C, Wang Y, Zhang L, Qiao L, Sun S, Shao M, Tang X, Ding P, Tang C, Cao Y, Zhou Y, Guo M, Wei R, Li N, Xiao Y, Duan J, Yang Y. AHSA1 ist ein vielversprechendes therapeutisches Ziel für die zelluläre Proliferation und Proteasom-Inhibitor-Resistenz beim multiplen Myelom. J Exp Clin Cancer Res. 2022 Jan 6;41(1):11. doi: 10.1186/s13046-021-02220-1. PMID: 34991674; PMCID: PMC8734095. (IF:11.161)
[16] Luo Q, Wu X, Zhao P, Nan Y, Chang W, Zhu X, Su D, Liu Z. OTUD1 aktiviert Caspase-unabhängige und Caspase-abhängige Apoptose durch Förderung der AIF-Kerntranslokation und des MCL1-Abbaus. Adv Sci (Weinh). 2021 Feb 8;8(8):2002874. doi: 10.1002/advs.202002874. PMID: 33898171; PMCID: PMC8061361. (IF:15.84)
[17] Luo Q, Wu X, Nan Y, Chang W, Zhao P, Zhang Y, Su D, Liu Z. TRIM32/USP11 gleicht die Stabilität von ARID1A und den onkogenen/tumorsuppressiven Status des Plattenepithelkarzinoms aus. Cell Rep. 2020 Jan 7;30(1):98-111.e5. doi: 10.1016/j.celrep.2019.12.017. PMID: 31914402. (IF:9.42)
[18] Sun X, Peng X, Cao Y, Zhou Y, Sun Y. ADNP fördert die neuronale Differenzierung durch Modulation der Wnt/β-Catenin-Signalisierung. Nat Commun. 12.06.2020;11(1):2984. doi: 10.1038/s41467-020-16799-0. PMID: 32533114; PMCID: PMC7293280. (IF:14.911)
[19] Yang X, Wang H, Xie E, Tang B, Mu Q, Song Z, Chen J, Wang F, Min J. Die Neuverdrahtung der ERBB3- und ERK-Signalgebung führt zu einer Resistenz gegen die FGFR1-Hemmung bei gastrointestinalem Krebs mit einer ERBB3-E928G-Mutation. Protein Cell. 2020 Dez;11(12):915-920. doi: 10.1007/s13238-020-00749-z. PMID: 32632529; PMCID: PMC7719122. (IF:14.872)
[20] Chen, T., Chen, Y., Chen, H. et al. Durch zwei Enzyme angetriebene, unbegrenzte DNA-Laufnanomaschine zur intrazellulären Bildgebung von schwach exprimierter microRNA. Nano Res. 12, 1055–1060 (2019). https://doi.org/10.1007/s12274-019-2344-5 (IF:8.21)
[21] Zhang X, Qi Z, Yin H, Yang G. Die Interaktion zwischen p53- und Ras-Signalisierung steuert die Cisplatin-Resistenz über HDAC4- und HIF-1α-vermittelte Regulation von Apoptose und Autophagie. Theranostik. 30. Januar 2019;9(4):1096-1114. doi: 10.7150/thno.29673. PMID: 30867818; PMCID: PMC6401400. (IF:8.12)
[22] Zou Y, Wang A, Shi M, Chen X, Liu R, Li T, Zhang C, Zhang Z, Zhu L, Ju Z, Loscalzo J, Yang Y, Zhao Y. Analyse von Redoxlandschaften und -dynamiken in lebenden Zellen und in vivo mithilfe genetisch kodierter Fluoreszenzsensoren. Nat Protoc. 2018 Okt;13(10):2362-2386. doi: 10.1038/s41596-018-0042-5. PMID: 30258175; PMCID: PMC6714056. (IF:13.49)
[23] Zhang K, Zhao X, Chen X, Wei Y, Du W, Wang Y, Liu L, Zhao W, Han Z, Kong D, Zhao Q, Guo Z, Han Z, Liu N, Ma F, Li Z. Verbesserte therapeutische Wirkung von aus mesenchymalen Stammzellen gewonnenen Exosomen mit einem injizierbaren Hydrogel zur Behandlung von Hintergliedmaßenischämie. ACS Appl Mater Interfaces. 12. September 2018;10(36):30081-30091. doi: 10.1021/acsami.8b08449. Epub 29. August 2018. PMID: 30118197. (IF:8.09)
[24] Hao H, Hu S, Chen H, Bu D, Zhu L, Xu C, Chu F, Huo X, Tang Y, Sun X, Ding BS, Liu DP, Hu S, Wang M. Verlust von endothelialem CXCR7 beeinträchtigt die vaskuläre Homöostase und die kardiale Umgestaltung nach Myokardinfarkt: Auswirkungen auf die Entdeckung kardiovaskulärer Medikamente. Circulation. 28. März 2017;135(13):1253-1264. doi: 10.1161/CIRCULATIONAHA.116.023027. Epub 2017 Feb 2. PMID: 28154007. (IF:18.881)
Zahlung und Sicherheit
Ihre Zahlungsinformationen werden sicher verarbeitet. Wir speichern weder Kreditkartendaten noch Zugriff auf Ihre Kreditkarteninformationen.
Anfrage
Sie können auch mögen
FAQ
Das Produkt ist nur für Forschungszwecke bestimmt und nicht für die therapeutische oder diagnostische Anwendung bei Menschen oder Tieren. Produkte und Inhalte sind durch Patente, Marken und Urheberrechte von
Für bestimmte Anwendungen sind möglicherweise zusätzliche geistige Eigentumsrechte Dritter erforderlich.