Nguyên liệu khuếch đại đẳng nhiệt chất lượng cao ——Giúp RT-LAMP nhạy hơn và nhanh hơn!
Công nghệ khuếch đại đẳng nhiệt có thể đạt được mục đích khuếch đại các chuỗi axit nucleic cụ thể trong điều kiện nhiệt độ không đổi. Do đặc điểm không cần thiết bị đặc biệt, thời gian khuếch đại ngắn và độ nhạy cao, nó đã cho thấy triển vọng ứng dụng tốt trong phát hiện tại chỗ và chẩn đoán tại chỗ và rất phù hợp để phát triển các công cụ chẩn đoán phân tử.
Trong bối cảnh đại dịch do virus corona mới gây ra với các chủng và khả năng lây nhiễm ngày càng gia tăng, công nghệ khuếch đại đẳng nhiệt có thể phát hiện virus corona mới nhanh chóng bằng các dụng cụ đơn giản, giúp giải quyết vấn đề một số bệnh nhân COVID-19 không thể xác nhận nhanh do thời gian phát hiện lâu và yêu cầu cao về thiết bị phát hiện và thuốc thử của các kỹ thuật PCR truyền thống. Vậy nguyên lý của RT-LAMP, một trong những kỹ thuật khuếch đại đẳng nhiệt là gì? Những nguyên liệu thô chất lượng cao nào
1. Nguyên lý của RT-LAMP là gì?
2. Thiết kế mồi cho RT-LAMP
3. Những nguyên liệu thô nào có thể
4. Hướng dẫn lựa chọn sản phẩm
1. Nguyên lý của RT-LAMP là gì?
Năm 2000, các học giả Nhật Bản Notomi và những người khác đã thành lập một công nghệ khuếch đại axit nucleic mới gọi là khuếch đại đẳng nhiệt qua vòng lặp (LAMP). LAMP sử dụng 4 đoạn mồi đặc hiệu được thiết kế cho 6 vùng của gen mục tiêu và sử dụng DNA polymerase dịch chuyển sợi để khuếch đại 109 bản sao của trình tự mục tiêu trong hàng chục phút trong điều kiện đẳng nhiệt (60~65 ℃). Kết quả được đánh giá bằng điện di gel agarose và kết quả dương tính cho thấy các dải hình thang. LAMP có các đặc điểm là độ đặc hiệu mạnh, độ nhạy cao, vận hành nhanh và đơn giản, chi phí thấp. Với sự cải tiến và hoàn thiện liên tục của công nghệ này, hiện nay nó được sử dụng để phát hiện nhiều loại vi sinh vật gây bệnh khác nhau.
RT-LAMP là phương pháp thêm phiên mã ngược vào hệ thống khuếch đại LAMP, có thể thực hiện phát hiện trực tiếp RNA virus. Hiệu suất khuếch đại của LAMP cực cao, do đó chỉ cần một lượng nhỏ cDNA cũng có thể khuếch đại một lượng lớn axit nucleic. Tiết kiệm thời gian phiên mã ngược trong quá trình khuếch đại axit nucleic, tốc độ phát hiện RNA được tăng tốc.
DNA ở trạng thái cân bằng động ở khoảng 65℃. Dưới tác động của DNA polymerase dịch chuyển sợi, bắt đầu từ đầu 3' của đoạn F2 của đoạn mồi FIP, nó được ghép nối với trình tự bổ sung của DNA khuôn mẫu để bắt đầu quá trình tổng hợp DNA dịch chuyển sợi. Đoạn mồi F3 bổ sung cho F3c ở đầu trước của F2c và lấy đầu 3' làm điểm bắt đầu để tổng hợp DNA của nó trong khi thay thế sợi DNA được tổng hợp bởi đoạn mồi FIP hàng đầu bằng tác động của DNA polymerase dịch chuyển sợi. Mở rộng về phía trước. Sợi DNA được tổng hợp bởi đoạn mồi F3 cuối cùng tạo thành một sợi đôi với một sợi DNA khuôn mẫu. Sợi DNA được tổng hợp bởi đoạn mồi FIP được thay thế bằng sợi mồi F3 để tạo thành một sợi đơn. Sợi đơn này có các đoạn F1c và F1 bổ sung ở đầu 5', do đó sự ghép cặp bazơ tự thân được thực hiện để tạo thành cấu trúc vòng. Và đoạn mồi BIP được lai hóa và kết hợp với mạch đơn, đầu 3' của đoạn mồi BIP được dùng làm điểm khởi đầu để tổng hợp mạch bổ sung, và cấu trúc được mở ra trong quá trình này.Sau đó, các đoạn mồi tương tự như F3 và B3 được chèn từ đoạn mồi BIP, quá trình ghép cặp bổ sung bazơ được thực hiện và một sợi bổ sung mới được tổng hợp từ đầu 3' làm điểm khởi đầu. Có các trình tự bổ sung ở cả hai đầu của DNA mạch đơn được thay thế và quá trình ghép cặp bazơ tự thân xảy ra để tạo thành cấu trúc vòng, do đó toàn bộ chuỗi có cấu trúc giống như quả tạ. Cấu trúc này là cấu trúc khởi đầu của chu kỳ khuếch đại của phương pháp RT-LAMP.
Trong cấu trúc hình quả tạ, quá trình kéo dài DNA được thực hiện bằng cách sử dụng đoạn F1 ở đầu 3' làm điểm bắt đầu và sử dụng chính nó làm khuôn mẫu. Và đoạn mồi FIP F2 lai với F2c mạch đơn trên vòng để bắt đầu một vòng phản ứng thay thế mạch mới. Axit nucleic mạch kép được tổng hợp từ đoạn F1 bị phân ly và tương tự như vậy, một cấu trúc vòng được hình thành trên axit nucleic. Có một dạng B2c mạch đơn trên cấu trúc vòng và B2 trên đoạn mồi BIP lai với nó để bắt đầu một vòng khuếch đại mới và một cấu trúc vòng được hình thành thông qua cùng một quá trình. Theo quá trình này, các trình tự bổ sung trên cùng một mạch tuần hoàn thông qua quá trình ghép cặp, kéo dài mạch và cuối cùng hình thành các cấu trúc có kích thước khác nhau.
2. Thiết kế mồi cho RT-LAMP
Thiết kế mồi là chìa khóa để khuếch đại và phát hiện RT-LAMP thành công. Mồi RT-LAMP bao gồm hai mồi ngoài (F3 và B3) và hai mồi trong (FIP và BIP). Mồi F3: Mồi ngoài thượng nguồn, bao gồm vùng F3, bổ sung cho vùng F3c của gen đích. Mồi FIP: mồi nội thượng nguồn, bao gồm vùng F2, vùng F2c ở đầu 3' của gen đích trong vùng F2 bổ sung cho vùng F1c ở đầu 5' của gen đích. Mồi BIP: mồi nội hạ nguồn, bao gồm vùng B2, vùng B2 bổ sung cho vùng B2c ở đầu 3' của gen đích và có cùng trình tự với vùng B1c ở đầu 5' của gen đích.
Tương tự như PCR, các nguyên tắc thiết kế mồi nên chú ý đến các yếu tố như thành phần bazơ, hàm lượng GC và cấu trúc phụ. Ngoài ra, cần lưu ý các điểm sau. Các phần đầu 5' của các mồi bên trong FIP và BIP, tức là F1c và B1c, thường có chiều dài 8-50 bp. Chiều dài này tốt nhất là từ 15 đến 25 bp và giá trị Tm lớn hơn các giá trị Tm của F2 và B2 ở phần đầu 3'. Các phần đầu 3' của các mồi bên trong FIP và BIP, tức là F2 và B2, thường có chiều dài 8-50 bp. Chiều dài này tốt nhất là từ 15-25 bp và giá trị Tm phù hợp với nhiệt độ tối ưu của DNA polymerase Bst được chọn trong thí nghiệm. Chiều dài của các mồi bên ngoài F3 và B3 thường có chiều dài 8-50 bp. Chiều dài này tốt nhất là từ 15-25 bp và giá trị Tm của nó nhỏ hơn giá trị của F2 và B2. Khi thiết kế mồi, cần xem xét cấu trúc vòng lặp và kích thước của trình tự mục tiêu. Khi số lượng bazơ trong vòng lặp lớn hơn 40bp và kích thước của trình tự mục tiêu là 130-200bp, hiệu suất khuếch đại là cao nhất.
3. Những nguyên liệu thô nào có thể Yeasen cung cấp?
3.1 [Nâng cấp mới] Yeasen Bst Cộng với DNA Polymerase
Mới được nâng cấp DNA Polymerase Hieff™ Bst Plus (Mã số: 14402ES, 14403ES) được thu được bằng cách biểu hiện và tinh chế gen DNA polymerase của Bacillus stearothermophilus sp thiếu miền exonuclease 5′→3′ ở E.coli.Khả năng thay thế chuỗi enzyme mạnh, có ưu điểm là độ nhạy cao, hiệu suất khuếch đại cao và khả năng chịu dUTP cao, có thể được sử dụng rộng rãi trong phát hiện mầm bệnh theo thời gian thực dựa trên công nghệ khuếch đại đẳng nhiệt.
3.1.1 Nhanh và độ nhạy cao
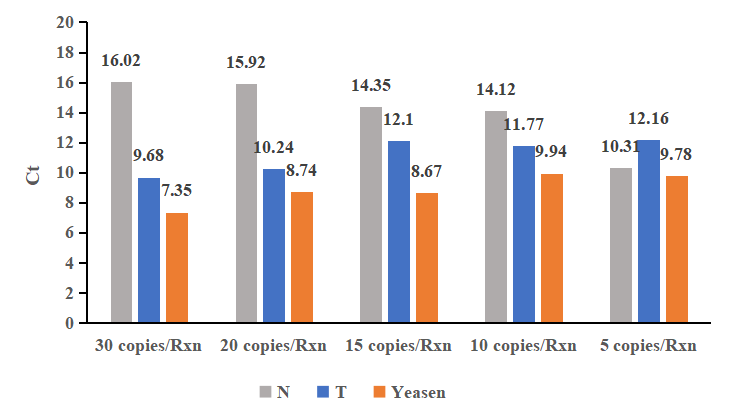
Hình 1. Phản ứng RT-LAMP được thực hiện với
4. Hướng dẫn lựa chọn sản phẩm
Các sản phẩm được cung cấp bởi
Bảng 1.Thông tin sản phẩm
| Định vị sản phẩm | Tên sản phẩm | Con mèo# |
| Enzym Bst có độ nhạy cao | 14402ES | |
| DNA Polymerase Hieff™ Bst Plus (2000 U/μL) (Hỏi thăm) | 14403ES | |
| Bộ hiển thị đèn RT-Lamp Phương pháp nhuộm huỳnh quang | 13762ES | |
| Phiên mã ngược thích hợp cho RT-Lamp | Phiên mã ngược Hifair™ Ⅲ (Hỏi thăm) | 11111ES |
| Hifair™ Ⅲ Reverse Transcriptase, không chứa Glycerol (Hỏi thăm) | 11297ES | |
| Chất ức chế RNase ở chuột | 10603ES | |
| 10703ES | ||
| UDG không bền với nhiệt | Uracil DNA Glycosylase (UDG/UNG), không bền với nhiệt, 1 U/μL (Hỏi thăm) | 10303ES |
| dUTP có độ tinh khiết cao | 10128ES |
Về việc đọc: