Genellikle yeni nesil dizileme (NGS) teknolojisi olarak adlandırılan yüksek verimli dizileme, Sanger dizilemesi gibi ilk DNA dizileme yöntemlerinden önemli bir sıçramayı temsil eder. NGS, milyonlarca olmasa da yüz binlerce nükleik asit molekül dizisinin eş zamanlı olarak profillenmesine olanak tanır. Avantajları arasında olağanüstü verimlilik, maliyet etkinliği, ölçeklenebilirlik ve geniş bir uygulama yelpazesi bulunur ve bu da onu dünya çapında baskın dizileme teknolojisi haline getirir.
NGS dizileme iş akışı dört temel aşamayı kapsar: örnek hazırlama, kütüphane oluşturma, dizileme ve veri analizi. Kütüphane oluşturmanın merkezinde, parçalanmış genomik DNA'nın her iki ucuna standartlaştırılmış NGS platform adaptör dizilerinin bağlanması yer alır. Bu adım, PCR amplifikasyonu yoluyla NGS cihazında dizilemeye hazırlanan bol miktarda kütüphane nükleik asit molekülü üretmeyi amaçlar. Örneğin doğasına bağlı olarak, NGS kütüphane oluşturma DNA kütüphane oluşturma ve RNA kütüphane oluşturma olarak kategorize edilebilir. Enzimler bu birbirine bağlı deneylerde önemli bir rol oynar. Peki, kütüphane oluşturma sürecinde hangi temel enzimler yer alır?
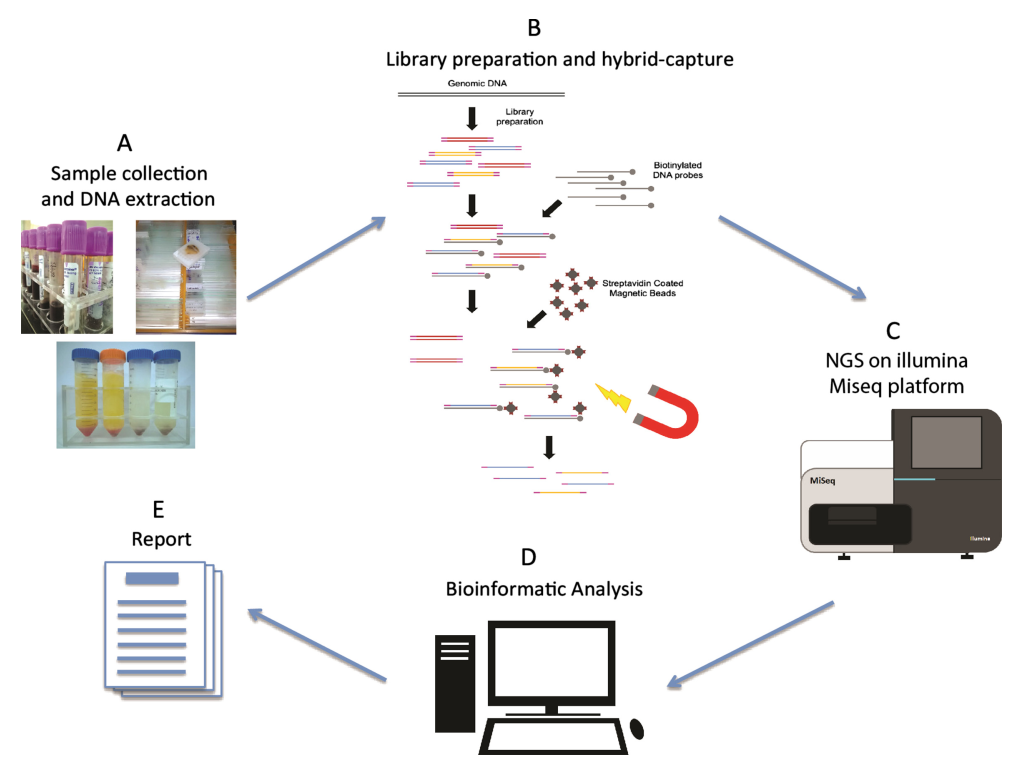
Şekil 1. Yeni nesil dizileme iş akışı[2]
1. DNA kütüphanesinin inşası ve temel enzimleri
2. RNA kütüphanesinin inşası ve temel enzimleri
3. DNA ve RNA kütüphanesi yapımında NGS çekirdek enzimleri için kılavuz
1. DNA kütüphanesinin inşası ve temel enzimleri
DNA kütüphanesi inşa sürecinde, TA klon ligasyon adaptörü kütüphanesi inşası şu anda en yaygın kullanılan teknoloji aracıdır. Ana kütüphane inşa süreci şu şekildedir:

Şekil 2. DNA kütüphanesi oluşturma süreci (Illumina)
1.1 DNA Parçalanması
Mevcut dizileyiciler genellikle 150-500 baz çifti (bp) aralığında bir dizileme uzunluğuna sahiptir. Sonuç olarak, büyük genomik DNA parçalarını daha küçük parçalara ayırmak için mekanik veya enzimatik parçalama yöntemleri kullanmak gerekli hale gelir. Mekanik parçalama nispeten yüksek örnek kaybına yol açabilir ve daha karmaşık bir operasyonel süreci içerir. Öte yandan, enzimatik sindirim genomik DNA'yı parçalamak için yaygın olarak kullanılan bir yöntemdir. Mekanik yöntemlerle karşılaştırıldığında, enzimatik sindirim daha uygun maliyetli ve basittir ve reaksiyon parçalama enziminin eklenmesinden sonra yalnızca belirli bir süre gerektirir.
Şu anda, kullanımda olan başlıca iki tür parça vardır. Biri, transpozon prensiplerine dayanan Tn5 transpozaza dayanırken, diğeri endonükleazların bir karışımını kullanır. Ancak, bu parçaların etkinliği, DNA'nın GC içeriği ve baz tercihlerinden etkilenebilir. Buna karşılık, Yeasen (Cat#12917) tarafından geliştirilen parçalar, kararlı bir sindirim etkisi sunar ve Tn5 transpozaza kıyasla önemli ölçüde daha düşük yer tercihi sergiler. FFPE örnekleri de dahil olmak üzere çeşitli DNA örnekleri için sürekli olarak mükemmel dizileme sonuçları verirler.
1.2 Son onarım, dA-Tailing
Parçalanmış DNA, 5'/3' yapışkan uçlar ve kör uçlu DNA üretecektir ve tüm yapışkan uçların kör uçlara dönüştürülmesi gerekir, buna 3' çıkıntılar kaldırılmalı ve 5' çıkıntılı DNA uçları doldurulmalıdır. Adaptör ligasyonu için TA ligasyonu kullanıldığında, DNA parçasının 5' ucunda fosforile edilmesi ve "T" yapışkan uçlu adaptöre tamamlayıcı olması için 3' ucuna "A" eklenmesi gerekir.Yukarıdaki işlem T4 DNA polimeraz, T4 polinükleotid kinaz ve T4 DNA polimerazının işbirliği ile tamamlanır. Tak DNA polimeraz.
T4 DNA polimeraz (Cat#12901) 5'→3' DNA polimeraz aktivitesine sahiptir, bu da 5'→3' yönünde DNA sentezini katalize edebilir ve 5' çıkıntılı ucu doldurabilir. Aynı zamanda, enzim ayrıca 3' çıkıntılı uçları kesmek için 3'→5' ekzonükleaz aktivitesine sahiptir, böylece yapışkan uçlar içeren DNA parçalarını kör uçlu DNA'ya dönüştürür.
Sentetik PCR primerlerinin ve adaptörlerinin 5' uçları genellikle fosfat grupları yerine hidroksil grupları olduğundan, T4 polinükleotid kinaz (Cat#12902), adaptör ligasyonunun bir sonraki adımına hazırlık olarak, ATP varlığında oligonükleotid zincirinin 5' -hidroksil ucuna ATP γ -fosfat gruplarının transferini katalize etmek için gereklidir.
S-Tak DNA polimeraz (Cat#13486) 5'→3' polimeraz aktivitesine sahiptir, bu da DNA'yı 5'→3' yönünden sentezleyebilir. Bu arada, PCR ürününün 3' ucuna bir nükleotid "A" ekleyebilen deoksinükleotidil transferaz aktivitesine sahiptir.

Şekil 3. Son onarım sürecinde birden fazla enzim yer alır

Şekil 4. S-taq, kılcal elektroforezle tespit edilen gen segmentlerinin 3' ucundaki ATCG'nin dört bazına A eklemede çok yüksek bir verimliliğe sahiptir.
1.3 Adaptör ligasyonu
Adaptörler, kütüphanenin önemli bir bileşenini oluşturur. Illumina dizileme alanında, yaygın olarak kullanılan Y tipi adaptörler P5/P7, Index ve Rd1/Rd2 SP dizilerini kapsar. Bunlar arasında, P5/P7 dizisi dizileme çipinde bulunan diziyle eşleşme amacına hizmet eder ve böylece analiz edilecek parçaları köprü amplifikasyonunu gerçekleştirmek için akış hücresine sabitler. Index dizisi, dizilemeye tabi tutulan karışık kütüphanedeki farklı örnekler arasında ayrım yapmak için kullanılırken, Rd1/Rd2 SP, Read1 ve Read2 dizileme primerlerine bağlanma bölgelerini belirtir.
Adaptör ligasyonu görevi için, T4 DNA ligazı (Cat#12996) standart seçimdir. Çift sarmallı DNA'da bulunan tek sarmallı çentikleri onarma ve bitişik nükleotidleri yeniden bağlama yeteneği gösterir.
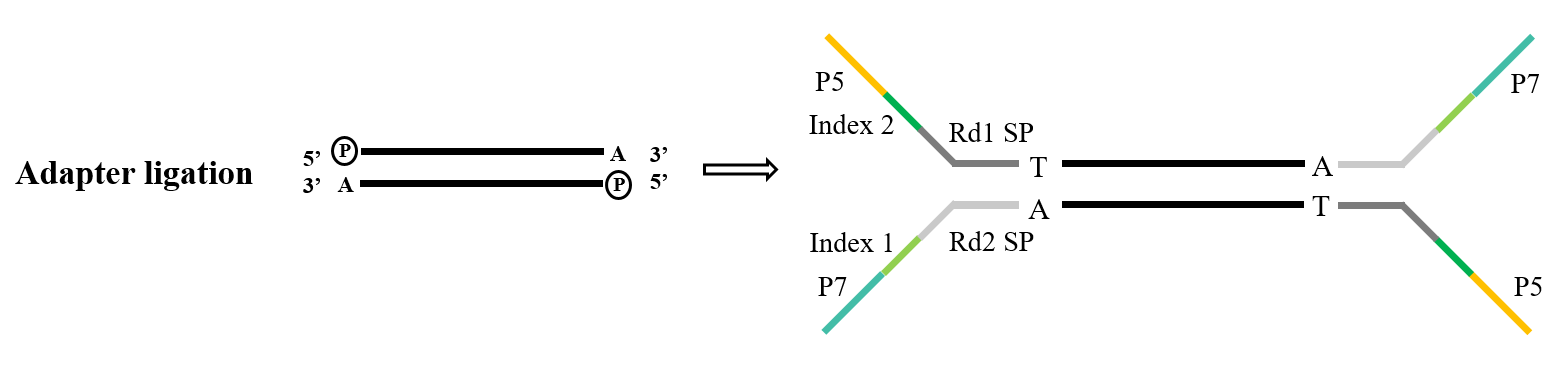
Şekil 5. Genel adaptör bağlama işlemi (Illumina)

Şekil 6. 170 bp DNA'nın 80 bp Adaptörlerle Bağlanmasıyla T4 DNA Ligaz Mutantlarının Doğrulanması.
1.4 PCR amplifikasyonu
PCR reaksiyonu yoluyla adaptörlerle yeterli DNA dizisi elde edin ve makinede örnek nükleik asit dizisinin dizilemesini tamamlayın. Hieff CanaceT.M. PCR'de yaygın olarak kullanılan Pro High-Fidelity DNA Polymerase (Cat#13476), 5'→3' polimeraz aktivitesine sahiptir ve DNA'yı 5'→3' yönünde sentezleyebilir. Ayrıca, amplifikasyon süreci sırasında bazların yanlış bir şekilde dahil edilmesini düzeltebilen ve DNA parçalarını hızla ve yüksek doğrulukla çoğaltabilen 3'→5' ekzonükleaz aktivitesine de sahiptir.
2. RNA kütüphanesinin inşası ve temel enzimleri
RNA türlerine göre RNA kütüphanesinin yapısı mRNA kütüphanesi, LncRNA kütüphanesi vb. olarak ayrılabilir. Geleneksel RNA kütüphanesi aşağıdaki süreçleri içerir:
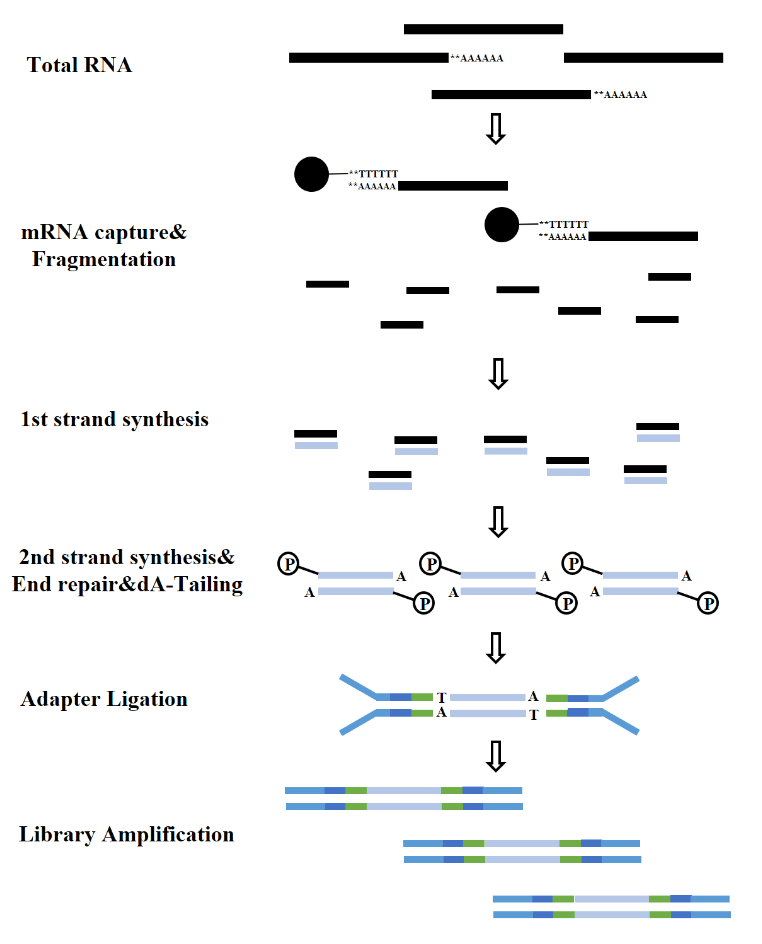
Şekil 7. mRNA kütüphanesi oluşturma süreci (Illumina)
2.1 RNA zenginleştirme
Ökaryotlar veya prokaryotlarla ilgili olsun, ribozomal RNA (rRNA) en bol bulunan RNA olarak öne çıkar ve toplam RNA içeriğinin %80'ine kadarını oluşturur. Bir örneğin toplam RNA'sını doğrudan dizilerken, dizileme verilerinin önemli bir kısmı rRNA ile ilişkili olacaktır. Bu müdahaleyi azaltmak için RNA zenginleştirme yöntemi kullanılmalıdır. Bunun için iki temel yöntem vardır: oligo-dT'ye dayalı mRNA zenginleştirme ve rRNA tükenme yöntemleri.
Ökaryotlarda, mRNA 3' ucunda belirgin bir poli(A) yapısı sergiler. Oligo-dT boncukları, örnekten transkripsiyona uğrayan tüm mRNA'yı yakalamak için kullanılabilir ve bu da onu özellikle yüksek kaliteli RNA örnekleriyle transkripsiyonel analiz için uygun hale getirir. Öte yandan, rRNA tükenme yöntemlerinin örnek kalitesi konusunda daha hoşgörülü gereksinimleri vardır ve hem düşük kaliteli örneklere (örneğin, FFPE örnekleri) hem de yüksek kaliteli RNA örneklerine ve prokaryotik örneklere uygulanabilir. Yaygın olarak kullanılan ticari yaklaşım, rRNA'yı çıkarmak için RNase H sindiriminin kullanılmasını içerir ve şu özel adımları izler:
- rRNA'ya bağlanmak üzere tasarlanmış spesifik oligonükleotid problarını sentezleyin.
- RNA-DNA hibrit zincirindeki RNA'yı parçalayabilen RNase H'yi (Kat#12906) kullanarak, problara bağlı rRNA'yı seçici olarak uzaklaştırın.
- Son olarak, DNA problarını sindirin DNase I (Kat#10325), hem tek hem de çift sarmallı DNA'yı parçalayabilen ve rRNA'yı etkili bir şekilde ortadan kaldıran. DNase I hakkında daha fazla bilgi için bu bağlantıyı takip edebilirsiniz.
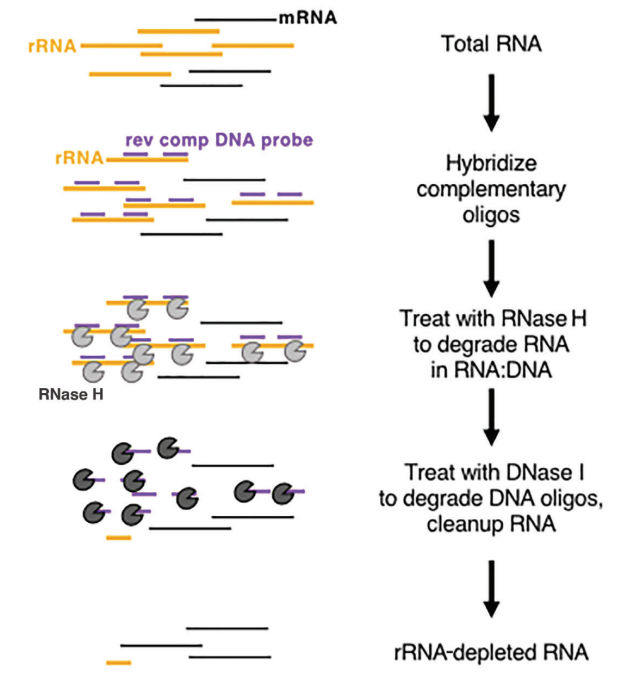
Şekil 8: Enzim bazlı rRNA tükenmesinin şematik diyagramı[5]
2.2 RNA Parçalanması
Genellikle iki değerlikli metal katyonların etkisi ve yüksek sıcaklık altında büyük RNA parçaları küçük parçalara ayrılır.
2.3 1. zincir cDNA sentezi
Elde edilen hedef RNA'nın cDNA'nın ilk ipliğine ters transkripsiyonu. RNA, ortamda bulunan RNazlar tarafından kolayca parçalandığından, RNase İnhibitörü (Kat#14672) ters transkripsiyon sırasında bu enzimlerin aktivitesini inhibe edebilir ve RNA'yı RNase bozunmasından koruyabilir. Aynı zamanda, ters transkriptaz (Cat#11112) şablon RNA'yı cDNA'ya ters transkribe etmek için kullanıldı. Ters transkriptaz, RNA'ya bağımlı DNA polimeraz aktivitesine sahiptir ve RNA'yı şablon olarak kullanarak 5'→3' yönünde bir cDNA sentezleyebilir. DNA'nın tek ipliği RNA şablonuna tamamlayıcıdır.
1. sırada iplik cDNA sentezi, aktinomisin D'nin dahil edilmesi, iplik özgül kütüphanelerin inşasını tartışmasız bir şekilde iyileştirerek zincir özgüllüğünü önemli ölçüde artırdı. Bu yenilik deneysel süreci hızlandırdı ve araştırmacılar için basitleştirdi.
Ancak, actinomycin D'nin dezavantajları da vardır: toksisite gösterir ve ışıktan korunmayı gerektirir. Günümüzde önceden karıştırılmış ve plak kütüphanesi oluşturma kitlerine olan talebin arttığı bir ortamda, ışığa karşı koruma gerekliliği plak kiti geliştirmelerinde sınırlamalar oluşturmaktadır.
Neyse ki, Yeasen ZymeEditor platformu, aktinomisin D'nin işlevini değiştiren çığır açıcı bir MMLV enzim mutantını (Inquiry) tanıttı. Yeni bir Kit (Kat: 12340ES), kokusuz, toksik olmayan vekaçınmaya gerek yok Hafiftir. Üstün zincir özgüllüğü sunarak sağlık ve ışık hassasiyetiyle ilgili endişeleri ortadan kaldırır.

Şekil 9: MMLV'nin, Standed RNA-seq'e katkıda bulunabilecek MMLV mutantlarını tanımlamak için mühendisliği
2.4 2. zincir cDNA sentezi
Ters transkripsiyonla üretilen tek zincirli cDNA oldukça kararsızdır ve DNA polimeraz I'in etkisi altında ikinci cDNA zincirinin derhal sentezlenmesini gerektirir. Bu ikinci zincir sentezi sırasında, RNA-DNA hibrit yapısından RNA zincirini çıkararak RNase H devreye girer. Şununla uyumlu çalışır: DNA polimeraz I (Cat#12903) cDNA'nın tamamlayıcı ikinci ipliğinin katalitik sentezini kolaylaştırmak için. DNA polimeraz I, 5'→3' DNA polimeraz aktivitesine sahiptir ve bir şablon ve primer tarafından yönlendirilerek, tek iplikli cDNA'yı 5'→3' yönünde tamamlayan bir dizi sentezler.
İşlemdeki sonraki adımlar, uç onarımı, dA-Tailing, adaptör ligasyonu ve PCR amplifikasyonunu içerir; bunların hepsi DNA kütüphanesi oluşturma prosedüründe ayrıntılı olarak açıklanmıştır ve burada tekrarlanmasına gerek yoktur. Ters transkripsiyon tamamlandıktan sonra, nükleik asit parçasının daha fazla parçalanmasına gerek olmadığını belirtmekte fayda var.
3. DNA ve RNA kütüphanesi yapımında NGS çekirdek enzimleri için kılavuz
Yeasen, üç büyük biyolojik reaktifin araştırma, geliştirme, üretim ve satışıyla uğraşan bir biyoteknoloji şirketidir: moleküller, proteinler ve hücreler. Yeasen Biotech şirketi, NGS kütüphane inşasıyla ilgili çeşitli enzimler üretmektedir. Aşağıdaki tablodan en uygun kütüphane inşa ürününü seçebilirsiniz.
Tablo 1.DNA ve RNA kütüphanesi yapımında NGS çekirdek enzimleri için kılavuz
| Tip | Ürün konumlandırma | Ürün adı | Kedi# |
| RNA kütüphanesi yapı | rRNA tükenme/2. iplik cDNA sentezi | 12906ES | |
| rRNA tükenme | 10325ES | ||
| 1. zincir cDNA sentezi | 14672ES | ||
| 11112ES | |||
| 2. zincir cDNA sentezi | 12903ES | ||
| RNA kütüphanesi yapı & DNA kütüphane yapı | Onarımı sonlandır | 12901ES | |
| 12902ES | |||
| dA-Tailing | 13486ES | ||
| Adaptör ligasyonu | 10301ES | ||
| PCR amplifikasyon | 2×Süper Canace® II Kütüphane Amplifikasyonu için Yüksek Sadakat Miks | 12621ES |
Tablo2.DNA ve RNA Kütüphanesi Hazırlık Seti
| İsim | Kedi# | Notlar | |
| DNA | Hieff NGS DNA Kütüphane Hazırlık Kiti | 13577ES | Tümör/ Mekanik yöntem |
| Hieff NGS OnePot Pro DNA Kütüphane Hazırlık Kiti V2 | 12194ES | Tümör/Enzimsel yöntem | |
| Yüksek NGS OnePot II Illumina için DNA Kütüphane Hazırlık Kiti | 13490ES | Pathgen/ Enzim/ normal süre (140dk) | |
| Hieff NGS OnePot Flash DNA Kütüphane Hazırlık Kiti | 12316ES | Pathgen/ Enzimsel/ Ultrahızlı (100dk) | |
| Hieff NGS DNA&RNA Kütüphanesi Ortak Hazırlama Kiti V2 | 12305ES | Pathgen/ Enzimetik/ DNA ve RNA Ortak Hazırlığı | |
| RNA | Hieff NGS Ultima Dual-mode mRNA Kütüphane Hazırlık Kiti | 12308ES | Oligo dT manyetik boncuklar olmadan, 11 tüp |
| Hieff NGS Ultima Dual-mode mRNA Kütüphane Hazırlık Kiti | 12309ES | oligo dT manyetik boncuklar artı, 14 tüp | |
| Hieff NGS® Ultima Dual-mod RNA Kütüphane Hazırlık Kiti | 12310ES | Önceden karıştırılmış versiyon, 5 tüp | |
| Hieff NGS ® EvoMax RNA Kütüphane Hazırlık Kiti (Önceden karıştırılmış versiyon) (aktinomisin D Özgür) | 12340ES | Önceden karıştırılmış versiyon, (Aktinomisin D Özgür) | |
| Hieff NGS® MaxUp rRNA Tükenme Kiti (Bitki) | 12254ES | Bitki | |
| Hieff NGS® MaxUp İnsan rRNA Tükenme Kiti (rRNA ve ITS/ETS) | 12257ES | İnsan |
Referanslar:
[1] Mardis, Elaine R. Yeni Nesil Dizileme Platformları[J]. Analitik Kimya Yıllık İncelemesi, 2013, 6(1):287-303.
[2] Gulilat M, Lamb T, Teft WA, ve diğerleri. Rezisyon tıbbı için bir araç olarak hedeflenen yeni nesil dizileme[J]. BMC Medical Genomics, 2019, 12(1):81.
[3] Lundberg KS, Dan DS, Adams M, ve diğerleri. Pyrococcus furiosus'tan izole edilen termostabil bir DNA polimeraz kullanılarak yüksek doğrulukta amplifikasyon[J]. Gene, 1991, 108(1):1-6.
[4] Miyazaki K. Endonükleaz V ile rastgele DNA parçalanması: DNA karıştırmaya uygulama[J]. Nükleik Asitler Araştırması, 2002, 30(24):e139.
[5] Baldwin A, Morris AR, Mukherjee N. RNA-seq için İnsan Ribozomal RNA'sını Tüketmek İçin Kolay, Maliyet Etkin ve Ölçeklenebilir Bir Yöntem[J]. Güncel Protokoller, 2021, 1(6):e176.